Transmission electron micrograph showing a mitochondrion (orange) inside a eukaryotic cell. The mitochondrion is enclosed by two membranes: an outer membrane and a highly folded inner membrane. These folds, called cristae, increase the surface area for energy production. Inside the inner membrane is the matrix, which contains enzymes for the citric acid cycle and mitochondrial DNA. This organelle is the primary site of ATP synthesis through oxidative phosphorylation, playing a central role in cellular energy metabolism.
Learning Objectives
Describe the three basic energy systems: CrP-ATP, glycolysis, and mitochondrial respiration (oxidative phosphorylation).
Explain the process of ATP formation and its importance in muscle contraction.
Identify the stages of carbohydrate metabolism and outline the key steps involved in glycolysis, conversion of pyruvate to acetyl-CoA, the TCA cycle, and the electron transport chain (ETC).
Compare and contrast glycolysis and glycogenolysis, including their ATP yields and regulatory enzymes.
Discuss the role of the phosphagen system in ATP production and its regulation by cellular constituents.
Understand the significance of the electron transport chain in oxidative phosphorylation and the production of ATP.
Explain the process of beta oxidation and its role in the metabolism of fatty acids to produce ATP.
Summarize the contribution of protein metabolism to ATP production and the conditions under which it becomes significant.
Analyze the relationship between carbohydrate, fat, and protein metabolism, highlighting the common intermediate, acetyl-CoA.
Evaluate the efficiency of different substrates (carbohydrates, fats, and proteins) in ATP production and their utilization during various intensities of exercise.
The Basic Energy Systems
The formation of adenosine triphosphate (ATP) is crucial for cellular energy storage and release. Cells have a limited capacity to store ATP, approximately 8 mmol/kg, necessitating its continuous synthesis. During muscle contraction, the demand for ATP can surge by up to 100-fold, depleting resting ATP levels within 2-3 seconds of intense exercise[1]. Consequently, skeletal muscle exhibits precise biochemical control over metabolic pathways through enzymatic regulation. This regulation ensures the production of ATP to replenish resting stores and support muscle contraction. Skeletal muscle cells can generate ATP via three metabolic pathways, either individually or in combination, as depicted in Figure 4.1. These pathways are collectively known as the energy systems. The three energy systems responsible for ATP production are:
1. CrP-ATP or the Phosphagen system
2. Glycolysis
3. Mitochondrial Respiration
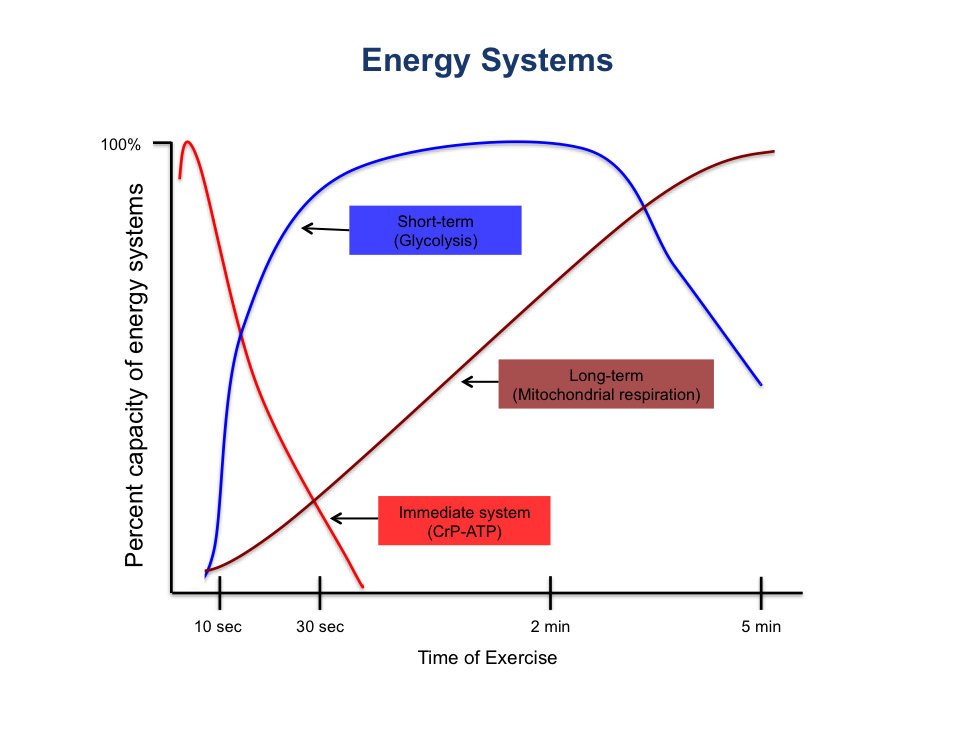
Figure 4.1 A line graph depicting the three basic energy systems (CrP-ATP, glycolysis, mitochondrial respiration) and the percent capacity of each system to generate ATP during 5 minutes of exercise.
The phosphagen energy system and glycolysis are metabolic pathways that can function without oxygen, collectively known as anaerobic metabolism. In contrast, mitochondrial respiration, also referred to as the oxidative system or oxidative phosphorylation, requires oxygen and constitutes aerobic metabolism. The energy required for exercise is derived from the interplay of both anaerobic and aerobic pathways. Typically, high-intensity, short-duration activities rely more heavily on anaerobic energy production, whereas prolonged, low to moderate-intensity activities depend on ATP generated from aerobic sources. It is important to note that all three energy systems operate continuously to maintain ATP levels, regardless of oxygen availability. Therefore, these terms are somewhat imprecise. The energy systems should be viewed as three engines that are always active, adjusting their output based on the demands placed upon them. One never exclusively uses “aerobic” or “anaerobic” metabolism but rather shifts reliance among the systems depending on exercise intensity and duration. These energy systems share a common central pathway, such as glycolysis, and function simultaneously.
CrP-ATP or the Phosphagen System
The phosphagen system is perhaps the simplest of the energy systems. It is the most immediate source of ATP and involves a series of coupled reactions to generate ATP. The phosphagen system operates by transferring phosphate groups to create ATP through two primary reactions: the creatine kinase reaction (CrP) and the adenylate kinase reaction. Cells store another high-energy phosphate molecule, creatine phosphate (CrP), which donates an inorganic phosphate (Pi) to adenosine diphosphate (ADP) to form ATP. Unlike ATP, CrP is not used directly for cellular work; its primary role is to regenerate ATP, ensuring a constant supply under resting conditions. Although the phosphagen system can regenerate ATP at high rates, the limited stores of CrP (approximately 26 mmol/kg wet weight) can be depleted in as little as 10 seconds[2].
The Creatine Kinase Reaction. The cellular store of creatine phosphate, also known as phosphocreatine (CrP), provides an almost immediate source of ATP. Reactions are often named after the enzyme that catalyzes them; in this case, the creation of ATP from CrP is facilitated by the enzyme creatine kinase, hence the term creatine kinase reaction. This reaction synthesizes ATP from CrP, with creatine kinase acting as the rate-limiting enzyme of the phosphagen system. At the onset of exercise and during the initial seconds, CrP is broken down to produce ATP and creatine from ADP and a proton (H+). Initially, this reaction alkalizes the cell by consuming a proton. A linear representation of the creatine kinase reaction is shown below:
Creatine phosphate + ADP ⇌ Creatine + ATP
During intense exercise, the creatine kinase reaction is driven to the right via the enzyme creatine kinase, resulting in the formation of ATP from creatine phosphate (CrP). In the body, this reaction consists of two coupled processes. The breakdown of CrP into creatine (Cr) and inorganic phosphate (Pi) is an exergonic reaction, which provides the energy needed for the endergonic synthesis of ATP from adenosine diphosphate (ADP), a proton (H+), and Pi. These coupled reactions help to delay acidosis during the initial seconds of intense exercise.
The Adenylate Kinase Reaction. Another high-energy phosphate molecule that can regenerate ATP is adenosine monophosphate (AMP). The adenylate kinase reaction, illustrated below, generates ATP from two ADP molecules with the enzyme adenylate kinase. During intense exercise, this reaction is also driven to the right, increasing the production of AMP. AMP acts as an allosteric activator for the enzyme phosphorylase (involved in glycogenolysis) and phosphofructokinase (involved in glycolysis), thereby enhancing carbohydrate catabolism. Phosphorylase and phosphofructokinase are crucial enzymes for carbohydrate metabolism and will be discussed in detail later in the chapter.
2 ADP ⇌ ATP + AMP
The AMP Deaminase Reaction. Under extreme acidic conditions, the purine nucleotide cycle facilitates the further breakdown of adenosine monophosphate (AMP) to help buffer acidosis with the enzyme, AMP deaminase. This process is known as the AMP deaminase reaction. In this reaction, AMP combines with water and a proton (H+) to produce inosine monophosphate (IMP) and ammonia (NH4). It is important to note that ammonia is toxic to both cells and the central nervous system. The ammonia generated by this reaction is transported into the bloodstream, where it is metabolized by the liver, excreted by the kidneys, or lost through sweat. This reaction, shown below, highlights the production of ammonia as a byproduct.
AMP + H2O + H+ → IMP + NH4
ATP Hydrolysis and Synthesis Reactions.The phosphagen system heavily relies on coupled reactions involving ATP, specifically ATP hydrolysis (ATPase reaction) and ATP synthesis (ATP synthetase reaction). ATP hydrolysis involves the breakdown of ATP with water to release energy for cellular activities. The enzyme ATPase catalyzes this reaction, splitting ATP into adenosine diphosphate (ADP), inorganic phosphate (Pi), a proton (H+), and free energy. This process is exergonic, meaning it releases free energy, which is then used for various cellular functions, such as muscle contraction. ATP hydrolysis and its products, highlighting the free energy utilized for cellular activities are shown below:
ATP + H₂O → ADP + Pi + energy + H+
The ATP hydrolysis reaction can also proceed in reverse, depending on the cell’s needs. In this case, ATP is generated, and water is produced as a byproduct. This process is known as ATP synthesis. ATP is synthesized from adenosine diphosphate (ADP), inorganic phosphate (Pi), a proton (H+), and energy, facilitated by the enzyme ATP synthetase or synthase. ATP synthesis is an endergonic reaction, meaning it requires activation energy to proceed. This process is illustrated:
ADP + Pi + energy + H+→ ATP + H2O
Regulation of the Phosphagen System.The phosphagen system is regulated by the concentration of cellular constituents. The presence of adenosine diphosphate (ADP) stimulates the phosphagen system, while the presence of cellular adenosine triphosphate (ATP) inhibits or prevents its reactions. The reactions within the phosphagen system are coupled, working together to create ATP during periods of high cellular demand and to store ATP as creatine phosphate (CrP) when ATP is abundant. The combined stores of ATP and CrP can sustain muscle energy needs for only 3 to 15 seconds during an all-out sprint. Beyond this duration, muscles must rely on other energy systems, such as glycolysis and mitochondrial respiration, for ATP generation[3].
Carbohydrate Metabolism
Glucose is a primary substrate for both exercise and resting metabolism. Typically, most carbohydrates used by skeletal muscle during exercise come from blood glucose. However, skeletal muscle can store up to 1 to 3 percent of its weight as glycogen, the storage form of glucose[4]. Upon complete oxidation, a single glucose molecule yields a net of 30 ATP. This complete oxidation occurs through the tricarboxylic acid (TCA) cycle and the electron transport chain. Understanding glucose metabolism is essential for exercise professionals, as it provides insights into the similarities between fat and protein metabolism. Carbohydrate metabolism consists of four stages:
Stage I: Glycolysis
Stage II: Conversion of pyruvate to acetyl-CoA
Stage III: TCA cycle
Stage IV: Electron transport chain (ETC) (Oxidative phosphorylation)
Stage I: Glycolysis
The phosphagen system has a limited capacity to generate ATP beyond the first 15 seconds of exercise. The second method of ATP production involves liberating energy from blood glucose, a process known as glycolysis. Glycolysis is an anaerobic metabolic pathway that does not require oxygen. Glycogen, the storage form of glucose in muscles, is broken down through a similar pathway called glycogenolysis.
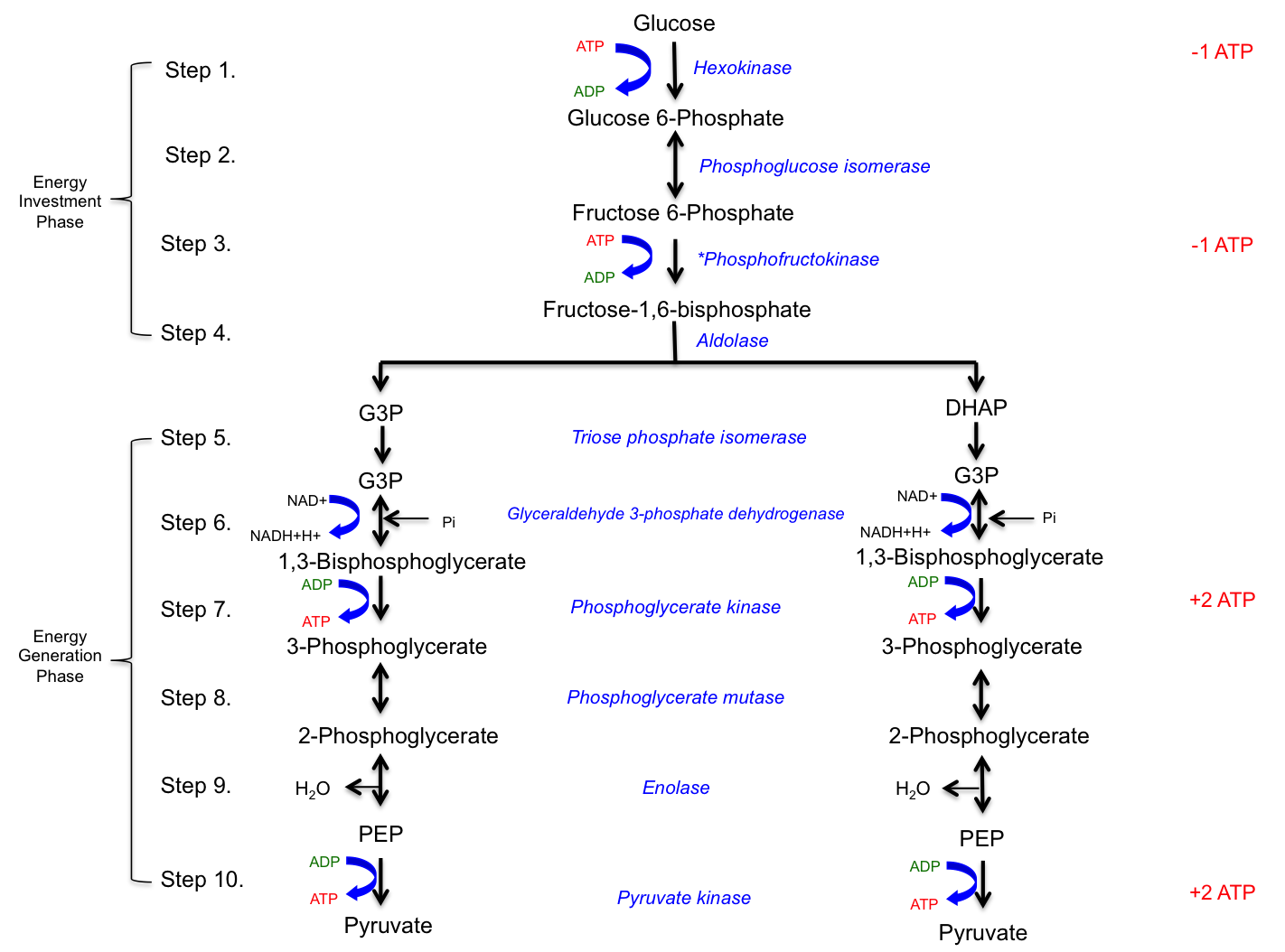
Glycolysis, which means the splitting of sugar (glucose), occurs in the cytoplasm of the cell. Glucose, with the chemical formula C6H12O6, is arranged in a 6-carbon ring structure and serves as the substrate for glycolysis. This pathway consists of 10 enzymatically catalyzed steps. The purpose of glycolysis is to break down the 6-carbon glucose molecule into two 3-carbon pyruvate molecules, which are then shuttled to the mitochondria for complete oxidation. The reactions in glycolysis can be divided into two phases:
Energy Investment Phase: The first four steps, where 2 ATP molecules are invested in steps 1 and 3 to provide the necessary activation energy.
Energy Generation Phase:Steps 5-10, where 4 ATP molecules are generated in steps 7 and 10 from a single glucose molecule.
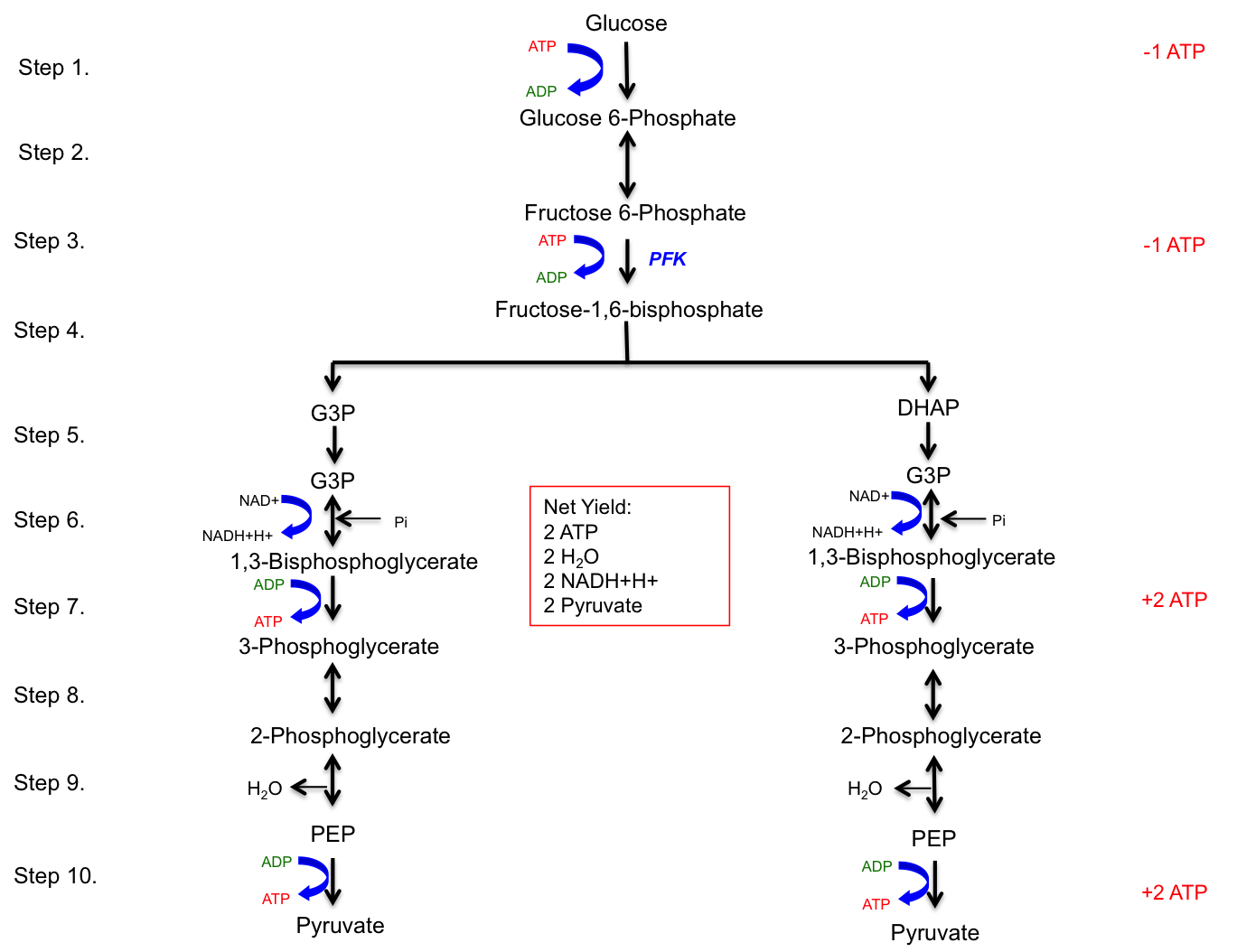
The net yield of glycolysis (Stage I of carbohydrate metabolism) includes 2 water H2O molecules, 2 pyruvate molecules, 2 ATP molecules, and 2 NADH+H+ molecules. The rate-limiting enzyme of glycolysis is phosphofructokinase (PFK), an allosteric enzyme that can change its conformation upon binding of effectors such as ATP or ADP. Glycolysis occurs in the sarcoplasm of muscle cells and provides a rapid means of generating ATP anaerobically. Each NADH+H+ produced in the cytoplasm results in the generation of 1.5 ATP equivalents.
Figure 4.2 Glycolysis. All enzymes are shown in blue text and the energy investment and generation phases are indicated with brackets. Tall of the ATP invested and generated is shown in red text on the right side of the pathway.
Table 4.1 The net yield of glycolysis (stage I of carbohydrate metabolism) from one glucose molecule.
The net yield of glycolysis
2 ATP
2 H2O
2 NADH+H+
2 Pyruvate
A simplified version of glycolysis is depicted in Figure 4.3, highlighting the major regulatory enzyme, phosphofructokinase (PFK), and the ATP tally. This figure provides a clear visual representation of the glycolytic pathway, emphasizing the key steps and the net production of ATP.
Figure 4.3 The ten steps of glycolysis and the net yield.
Glycogenolysis
Glycogenolysis is the metabolic pathway in which stored muscle glycogen is used as a substrate instead of blood glucose. This pathway involves the breakdown of glycogen into pyruvate and is very similar to glycolysis. However, the first step of glycogenolysis does not require an investment of ATP. Instead, glycogen is converted to glucose 1-phosphate by the enzyme phosphorylase. Phosphorylase adds an inorganic phosphate to the carbon structure, which is then converted to glucose 6-phosphate. From this point, the metabolic pathway follows the same steps as glycolysis. Consequently, the complete oxidation of glycogen yields a net ATP production of 31 ATP. Figure 4.4 illustrates the steps in glycogenolysis that differ from glycolysis.
Figure 4.4 The first two steps of glycogenolysis differ from glycolysis but are the same following conversion of the substrates to glucose 6-phosphate.
Comparison of Glycogenolysis and Glycolysis. Figure 4.5 provides a comparison of glycogenolysis and glycolysis. One key difference is that glycogenolysis does not require an ATP investment in the first step. As a result, the net ATP yield from the complete oxidation of glycogen is 31 ATP, compared to the 30 ATP yield from glycolysis. This comparison highlights the efficiency of glycogenolysis in energy production.
Figure 4.5 Comparison of glycogenolysis and glycolysis. Glycogenolysis does not invest 1 ATP during step 1 and 2. Glucose 1-phosphate is converted to Glucose 6-phosphate and then the following steps are the same as the glycolytic pathway. Thus, glycolysis has an ATP yield of 30 ATP and glycogenolysis has an ATP yield of 31.
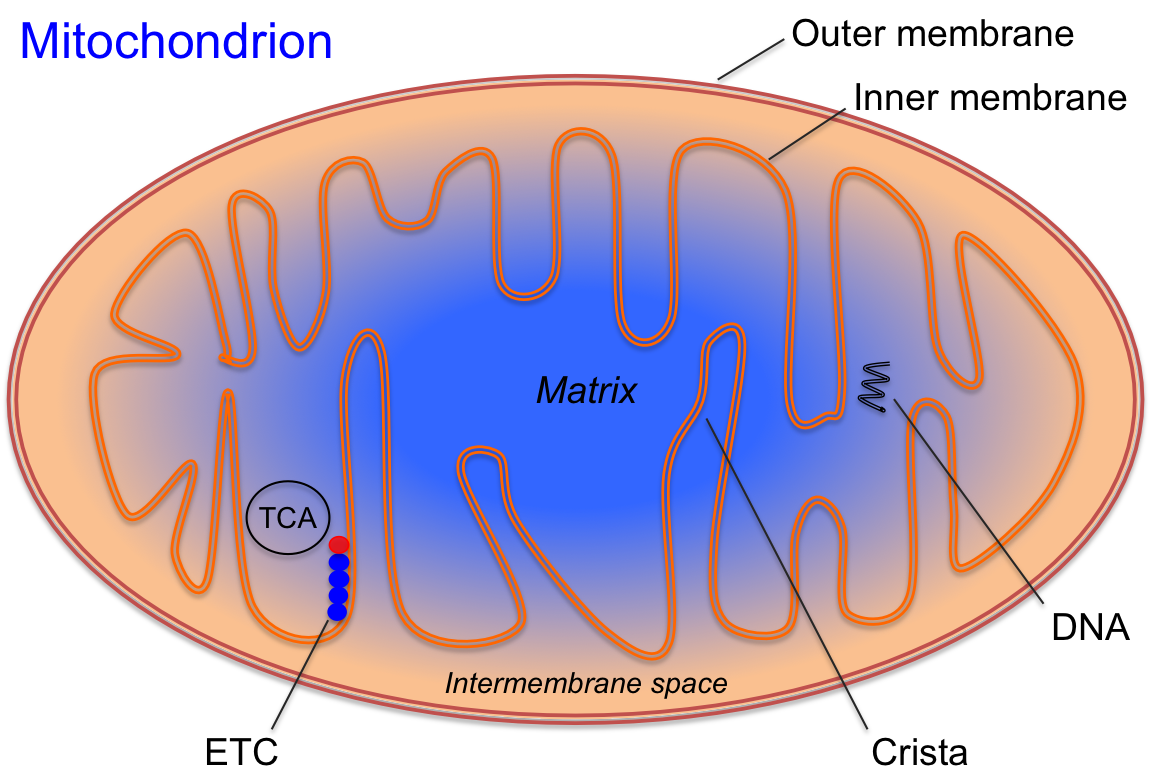
Mitochondria, illustrated in Figure 4.6, are often referred to as the “powerhouses” of the cell because they are the primary site of ATP production. Mitochondria are distributed throughout the cytoplasm of cells, with their number per cell ranging from fewer than a hundred to several thousand, depending on the cell’s energy requirements[5]. Mitochondria can also vary in size and shape. The basic structure of a mitochondrion, shown in Figure 4.11, consists of two lipid bilayer-protein membranes: an outer membrane and an inner membrane. The space between these membranes, known as the intermembrane space, is crucial for creating a proton gradient during the electron transport chain (ETC).
Figure 4.6 The mitochondrion is an organelle inside of the cell and is where mitochondrial respiration (oxidative phosphorylation) occurs.
The inner membrane is characterized by folds called cristae, which extend into the mitochondrial matrix. The matrix, the inner cavity of the mitochondrion, contains a high concentration of enzymes responsible for nutrient oxidation, such as those involved in the tricarboxylic acid (TCA) cycle. Additionally, mitochondria possess their own DNA, enabling them to self-replicate and produce proteins as needed to meet the cell’s ATP demands.
Several metabolic pathways within the mitochondria complete the oxidation of substrates. This chapter will discuss three key metabolic processes that occur within the mitochondria:
1) Conversion of Pyruvate to Acetyl-CoA
2) TCA Cycle
3) Electron Transport Chain (ETC)
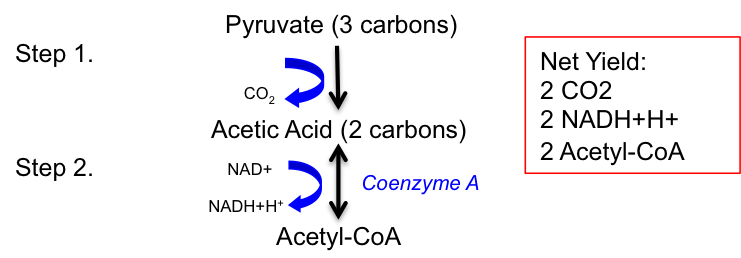
Stage II: Conversion of Pyruvate to Acetyl-CoA
To complete the oxidation of a single glucose molecule, the two pyruvate molecules formed during glycolysis must undergo further catabolism. This process converts pyruvate into acetyl-coenzyme A (acetyl-CoA), often referred to as the intermediate molecule of metabolism because it is a common product of the catabolism of proteins, fats, and carbohydrates. Pyruvate molecules produced in glycolysis are transported into the mitochondria. Once inside the mitochondrial matrix, pyruvate is converted into acetic acid, releasing a carbon dioxide (CO2) molecule as a byproduct.
The conversion of pyruvate to acetyl-CoA (Stage II of carbohydrate metabolism) occurs within the mitochondria and involves a two-step process, as shown in Figure 4.7. In the first step, pyruvate (a 3-carbon molecule) is converted into acetic acid (a 2-carbon molecule), releasing CO2. In the second step, acetic acid is converted into acetyl-CoA by coenzyme A (CoA). During this reaction, NAD+ oxidizes acetic acid by removing hydrogen, resulting in the reduction of NAD+ to NADH+H+.
The net yield from the conversion of pyruvate to acetyl-CoA (Stage II of carbohydrate metabolism) from a single glucose molecule includes 2 CO2, 2 NADH+H+, and 2 acetyl-CoA molecules, as summarized in Table 4.2.
Figure 4.7 Conversion of pyruvate to acetyl-CoA and the net yield.
Table 4.2 The net yield of conversion of pyruvate to acetyl Co-A (stage II of carbohydrate metabolism) from 1 glucose molecule.
The net yield of conversion of pyruvate to
acetyl-CoA
2 CO2
2 NADH+H+
2 Acetyl-CoA
Stage III: TCA Cycle
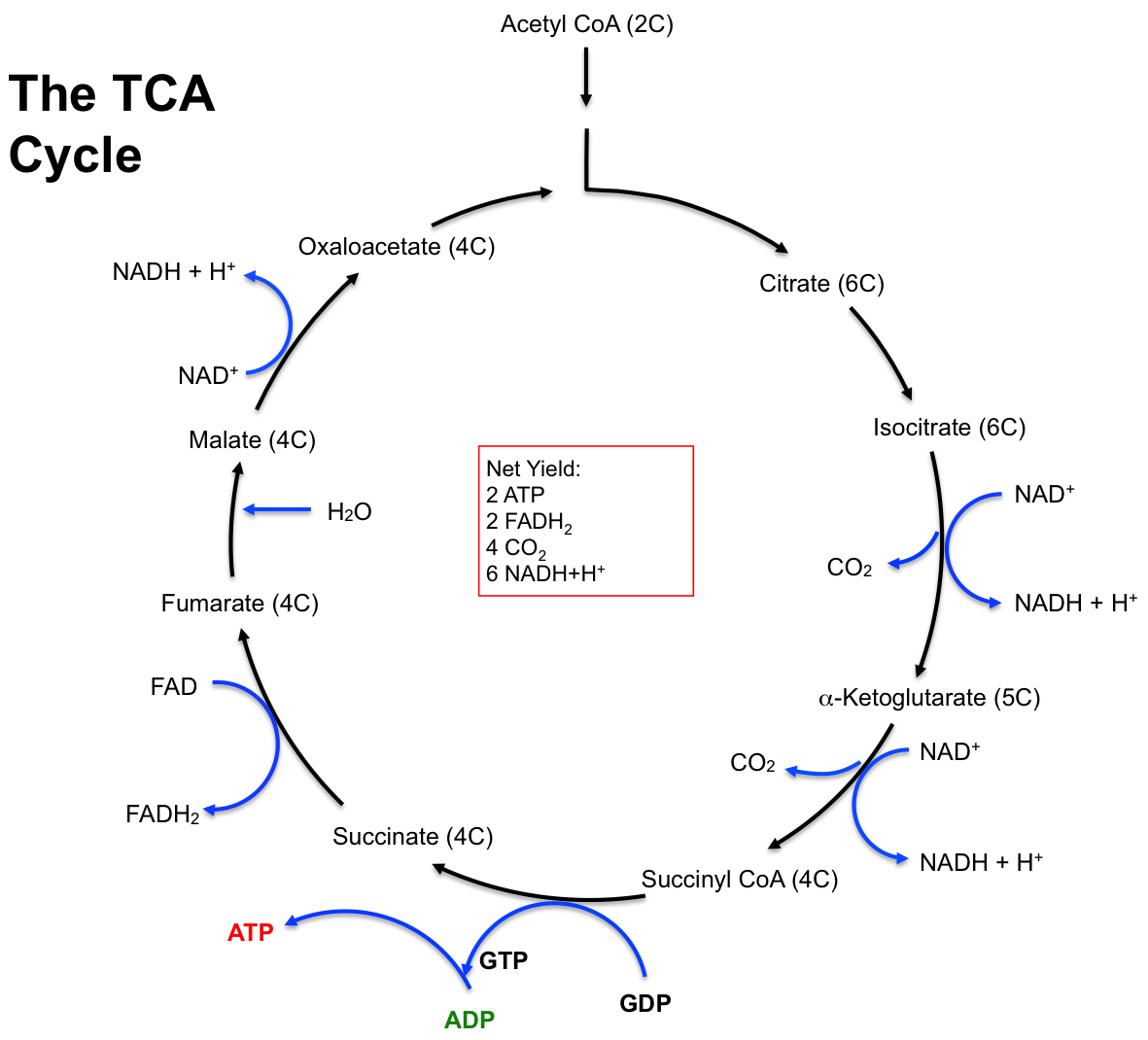
The tricarboxylic acid (TCA) cycle, also known as the citric acid cycle or Krebs cycle, is a series of enzyme-catalyzed chemical reactions that are essential for aerobic respiration. This cycle completes the oxidation of carbohydrates, fats, and proteins within the mitochondrial matrix. Named after Hans Krebs, the 1953 Nobel Prize recipient for his research on these reactions, the TCA cycle is crucial for the complete oxidation of glucose, which does not occur until acetyl-CoA is degraded to carbon dioxide (CO2) and hydrogen atoms. The hydrogen atoms released from acetyl-CoA are subsequently oxidized in the electron transport chain (ETC), releasing significant amounts of energy to form ATP[6].
Entry into the TCA cycle requires the breakdown of carbohydrates, fats, or proteins into acetyl-CoA. Focusing on the formation of acetyl-CoA from pyruvate, the TCA cycle involves eight steps, beginning with acetyl-CoA (a two-carbon molecule) and ending with the regeneration of oxaloacetate (a four-carbon molecule). Initially, acetyl-CoA combines with oxaloacetate to form citrate (a six-carbon molecule). This is followed by a series of reactions that regenerate oxaloacetate and produce two molecules of CO2. Due to the cyclic nature of the TCA cycle, oxaloacetate is not a final product but is continuously combined with acetyl-CoA to restart the cycle (Figure 4.8).
Since two acetyl-CoA molecules are formed from one glucose molecule during Stage II of carbohydrate metabolism, this results in two turns of the TCA cycle. Each turn of the TCA cycle yields one ATP (via GTP), three NADH+H+, one FADH2, and two CO2molecules. Therefore, the net yield from one glucose molecule (Table 4.3) is double the yield of a single turn. Each NADH+H+ produced in the TCA cycle will yield 2.5 ATP equivalents in the ETC, and each FADH2 will yield 1.5 ATP equivalents. The rate-limiting enzyme of the TCA cycle is isocitrate dehydrogenase.
Figure 4.8 The reactions and the net yield of the TCA cycle, also known as the Krebs cycle and the Citric Acid cycle.
Table 4.3 The net yield of the TCA cycle (stage III of carbohydrate metabolism) from one glucose molecule.
Net Yield of the TCA cycle (remember: 2 acetyl-CoA molecules)
2 ATP (by way of GTP)
6 NADH+H+
2 FADH2
4 CO2
As discussed in Chapter 3, hydrogen atoms consist of a proton and an electron. When hydrogen is transferred to NAD+ and FAD, these electron carriers also transport protons. Following the TCA cycle, a single glucose molecule is completely oxidized through a series of redox reactions, where NAD+ and FAD are reduced to NADH+H+ and FADH2, respectively. The NADH+H+ and FADH2 produced in Stages II and III of carbohydrate metabolism shuttle their hydrogens to the electron transport chain (ETC), where they are oxidized back to NAD+ and FAD. Both NADH+H+ and FADH2 return to their oxidized forms by releasing electrons to the electron carriers within the ETC.
The origin of NADH+H+ determines the number of ATP equivalents produced during the complete oxidation of glucose. Each NADH+H+ created in the cytoplasm yields 1.5 ATP equivalents, while NADH+H+ produced inside the mitochondria yields 2.5 ATP equivalents. Additionally, each FADH2 molecule produced in the mitochondria yields 1.5 ATP equivalents. The difference in ATP yield is due to the double membrane structure of the mitochondria. NADH+H+ created in the cytoplasm must transfer its hydrogen into the mitochondria because it cannot cross the double membrane barrier. This transfer is facilitated by a “shuttle” mechanism that transfers hydrogen from NADH+H+ to a FAD molecule inside the mitochondria, reducing FAD to FADH2, which then carries the hydrogen to the ETC. Consequently, NADH+H+ created in the cytoplasm has the same ATP yield (1.5 ATP) as FADH2.
The mechanism by which hydrogen carried by NADH+H+ and FADH2 is used to generate ATP will be further explained in the ETC section. The total ATP produced from one glucose molecule during carbohydrate metabolism, including ATP equivalents, is summarized and organized by stage in Table 4.4.
Table 4.4 Total ATP produced from one glucose molecule during carbohydrate metabolism.
Total ATP production from one glucose molecule (net yields)
ATP yield
Stage I. Glycolysis:
ATP
2 NADH+H+ ⇒ 2 FADH2 (to ETC)
2
3
Stage II. Conversion of pyruvate to acetyl CoA:
2 NADH+H+ (to ETC)
5
Stage III. TCA cycle (remember: 2 cycles):
ATP (at one site)
NADH+H+ at three steps (to ETC)
FADH2 at one step (to ETC)
2
15
3
Total ATP from one molecule of glucose:
30 ATP
Stage IV: Electron Transport Chain (Oxidative Phosphorylation)
The aerobic production of ATP, known as oxidative phosphorylation, occurs within the mitochondria. The pathway responsible for this process is the electron transport chain (ETC), where the majority of ATP is synthesized. The theory of the electron transport chain was proposed by British physiologist Peter Mitchell in 1961 but was not widely accepted until 1978. The mechanism explaining the aerobic formation of ATP is known as the chemiosmotic hypothesis. This hypothesis describes the transfer of electrons along a chain of proteins, releasing energy used to “pump” protons through the inner mitochondrial membrane. This creates an ion gradient and potential energy within the intermembrane space, which can then be harnessed to recombine inorganic phosphate (Pi) with adenosine diphosphate (ADP) to form ATP. At the end of the chain, oxygen combines with the electrons and two protons to form water, which is why oxygen is essential as the final electron acceptor in the ETC.
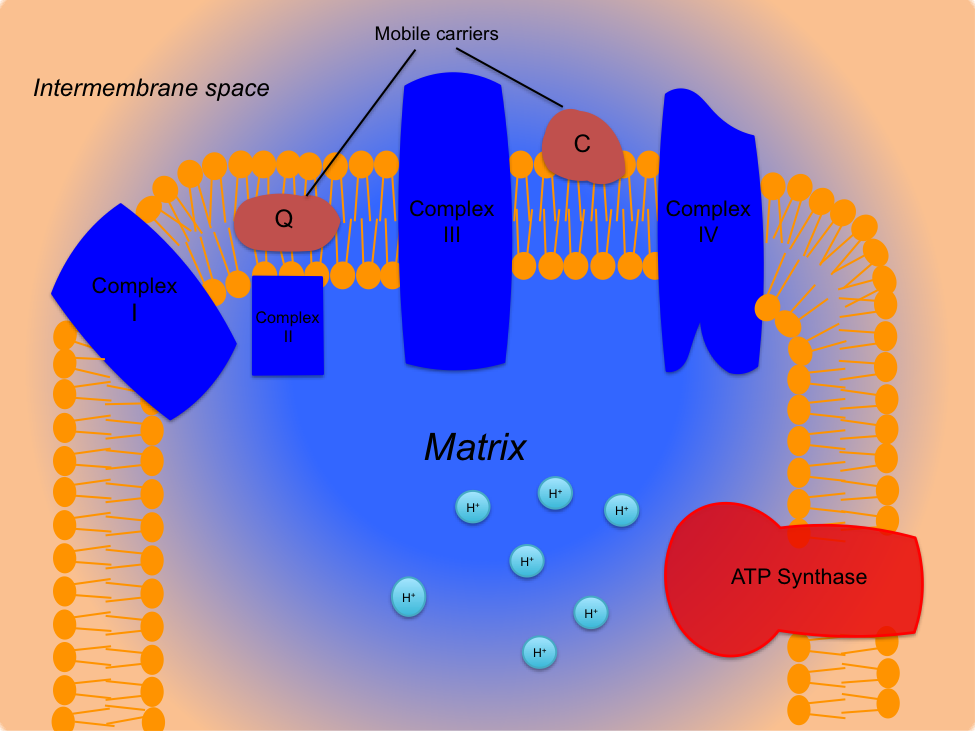
Oxidative phosphorylation results from a complex interaction between the TCA cycle and the ETC. The electron carriers NADH+H+ and FADH2 produced in these pathways play a crucial role in ATP synthesis within the ETC. The ETC is located in the inner mitochondrial membrane and consists of four protein complexes, known as the cytochrome chain, two mobile carriers, and ATP synthase. The ETC is illustrated in Figure 4.9. The cytochrome chain includes transmembrane proteins named complex I (NADH dehydrogenase), complex II (succinate dehydrogenase), complex III (cytochrome c reductase), and complex IV (cytochrome c oxidase). The mobile carriers are coenzyme Q10 (mobile carrier Q) and the cytochrome c complex (mobile carrier C). Finally, the enzyme ATP synthase (also known as ATP synthetase) is located at the end of the cytochrome chain and catalyzes the formation of ATP from ADP and Pi. Cytochrome oxidase (complex IV) is the rate-limiting enzyme of the ETC.
Figure 4.9 The electron transport chain is made up of four protein complexes; complex I (NADH dehydrogenase), complex II (succinate dehydrogenase), complex III (cytochrome c reductase), complex IV (cytochrome c oxidase): two mobile carriers; Q (coenzyme Q10) and C (cytochrome c complex): and ATP synthase. Protons (H+) are represented as blue circles.
The electron transport chain (ETC) involves a series of redox reactions that generate potential energy to synthesize ATP. The process begins with the arrival of NADH+H+ from Stage II (conversion of pyruvate to acetyl-CoA) or Stage III (TCA cycle) of carbohydrate metabolism. NADH+H+ donates two electrons to Complex I of the ETC and is oxidized to NAD+, with its protons being deposited into the matrix. As electrons enter Complex I, a redox reaction occurs, creating a pump that translocates four protons from the matrix into the intermembrane space of the mitochondria.
The electrons are then transferred to mobile carrier Q, and NAD+ returns to its original source to pick up more hydrogen. The electrons move from mobile carrier Q to Complex III, where another redox reaction occurs, pumping four more protons into the intermembrane space. The electrons are then shuttled from Complex III to Complex IV by mobile carrier C. Upon reaching Complex IV, a redox reaction pumps two additional protons into the intermembrane space. Complex IV, now in its reduced form, transfers the electrons to oxygen, the final electron acceptor. Oxygen combines with the electrons and two protons from the matrix to form water (H2O), known as “metabolic water,” which accounts for 10-20% of total daily fluid intake. Using NADH+H+ as the electron donor results in ten protons being pumped into the intermembrane space.
A similar process occurs with FADH2 as the electron donor. FADH2 donates two electrons to Complex II, is oxidized to FAD, and releases its protons into the matrix. Complex II passes the electrons to mobile carrier Q, continuing down the ETC towards Complex IV. This process bypasses Complex I, resulting in only six protons being pumped through the chain.
The protons in the intermembrane space create an electrochemical gradient, forming potential energy. This gradient, with a higher proton concentration in the intermembrane space than in the matrix, activates ATP synthase. Protons flow through ATP synthase, causing a rotary action that generates enough free energy to phosphorylate ADP, synthesizing ATP. For every four protons that flow through ATP synthase, one ATP is created. Therefore, each NADH+H+ produced in the mitochondria yields 2.5 ATP, and each FADH2 yields 1.5 ATP[7].
In summary, the ETC is where the greatest amount of ATP is synthesized. It involves the oxidation of NADH+H+ and FADH2 and the phosphorylation of ADP by ATP synthase, facilitating oxidative phosphorylation and aerobic metabolism. Table 4.4 summarizes the ATP yield from one glucose molecule.
Beta Oxidation (Oxidation of Fatty Acids)
Fats are also utilized in mitochondrial respiration to create ATP during exercise. Muscle and liver glycogen stores provide approximately 2,500 kcal of energy, while fat stored in muscle fibers and fat cells can supply 70,000 to 75,000 kcal, even in a lean adult[8]. Fats are stored as triglycerides and broken down by the enzyme hormone-sensitive lipase into free fatty acids and glycerol, a process called lipolysis[9]. Free fatty acids released into the blood can enter muscle fibers for oxidation. A triglyceride consists of one glycerol molecule and three fatty acids, with most triglycerides stored in adipose tissue. Limited quantities are stored in muscle cells, providing an intramuscular source of free fatty acids.
Fat is an excellent storage fuel because it is stored dry, without excess water, unlike glycogen, which is diluted with water. Fat yields about 9.13 kcal/g and contains many oxidizable carbons and hydrogen. Fat stores are substantial compared to glycogen and can last for weeks, even with heavy exercise.
Fatty acids, composed of long carbon chains, must be broken down into acetyl-CoA through beta oxidation to be used as fuel. Beta oxidation involves a series of reactions that break down fatty acids into pairs of carbons, forming acetyl-CoA and hydrogen protons. Acetyl-CoA enters the TCA cycle, and the hydrogen atoms are carried by NADH+H+ and FADH2 to the ETC. Each acetyl-CoA produced by beta oxidation generates 3 NADH+H+, 1 FADH2, and 1 ATP (from GTP) per acetyl-CoA molecule.
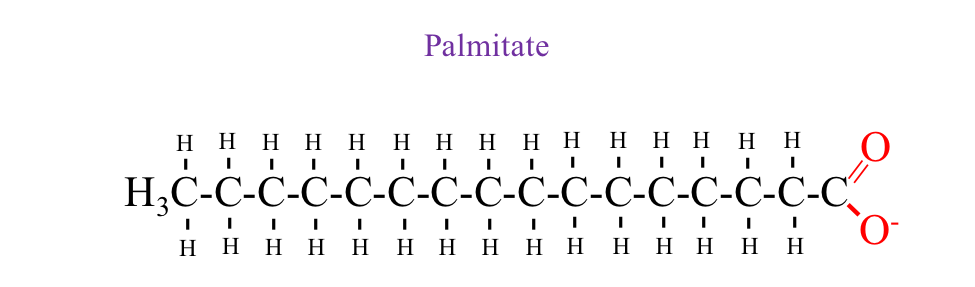
Figure 4.10 Palmitate is a 16-Carbon fatty acid chain commonly used in metabolism. The molecule consists of a straight hydrocarbon chain with 15 methylene (–CH₂–) groups and a terminal carboxyl group (–COOH). Palmitate is a key substrate in β-oxidation, where it is broken down in the mitochondria to generate acetyl-CoA, NADH+H+, and FADH₂, contributing to ATP production through the TCA cycle and oxidative phosphorylation.
ATP Production from Fatty Acids.There are various forms of fat, and the amount of ATP produced depends on the specific fatty acid being oxidized. For example, consider palmitate, a 16-carbon fatty acid chain illustrated in Figure 4.10. Theoretically, 106 ATP molecules can be produced from the oxidation of one palmitate molecule. During beta oxidation, palmitate is cleaved seven times to yield eight acetyl-CoA molecules. A helpful equation shown below shows how the number of cleavages required for any fatty acid length is mathematically expressed:
Number of times to cleave = (number of carbons / 2) – 1
To initiate beta oxidation, 2 ATP molecules are required to activate the fatty acid (note: 1 ATP is used for activation, but since it is hydrolyzed to AMP, this is equivalent to 2 ATP). Once activated, the fatty acid does not need to be reactivated. The first reaction, following the metabolism of acetyl-CoA in the TCA cycle, yields 12 ATP. The remaining acetyl-CoA molecules yield 14 ATP each until the last four carbons of the fatty acid chain. The last four carbons yield 4 ATP from beta oxidation and 10 ATP from each acetyl-CoA oxidized in the TCA cycle, totaling 24 ATP. The carboxylate ending of the palmitate molecule decreases the ATP yield of the last four carbons. In total, the complete oxidation of one 16-carbon palmitate molecule yields approximately 106 ATP. This demonstrates the significant capacity of fat oxidation for ATP synthesis compared to carbohydrate and protein oxidation[10].
The advantage of having more carbon and hydrogen atoms in free fatty acids than in glucose is that more acetyl-CoA is formed from fat metabolism, providing more hydrogens for the ETC. Although fat provides more kilocalories of energy per gram than carbohydrate, fat oxidation requires more oxygen. Oxygen delivery is limited by the oxygen transport system, making carbohydrate the preferred substrate during high-intensity exercise[11]. During high-intensity exercise, the maximum rate of ATP production from fat oxidation is insufficient to match ATP utilization, explaining the reduction in an athlete’s pace when carbohydrate stores are depleted, and fat becomes the primary fuel source[12].
Protein Oxidation (Metabolism)
Protein is not a major fuel source during exercise but can be broken down into amino acids through various metabolic processes. These amino acids are converted into glucose (gluconeogenesis), pyruvate, acetyl-CoA, or TCA cycle intermediates. Proteins contribute 3% to 18% of energy requirements during prolonged activity, but their contribution to ATP production is minimal during short-term exercise[13]. The major amino acids oxidized in skeletal muscle are branched-chain amino acids (leucine, isoleucine, and valine), along with alanine, aspartate, and glutamate[14]. When degraded, amino acids are eliminated through the formation of urea and ammonia. Ammonia is toxic to cells and is associated with muscle fatigue[15].
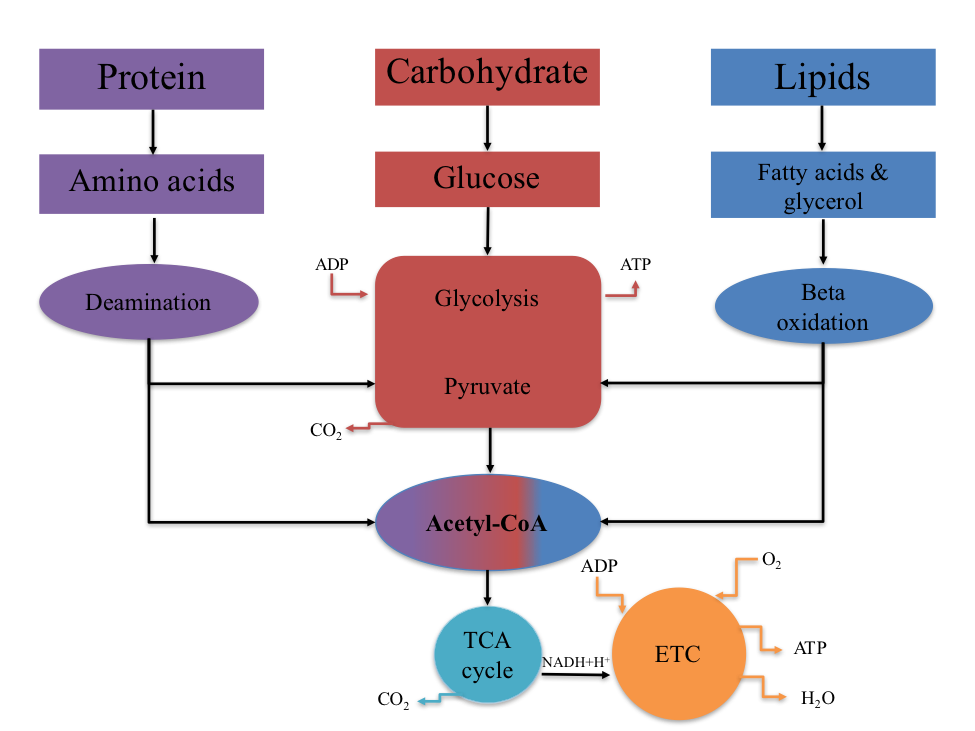
The relationship between protein, carbohydrate, and fat metabolism is illustrated in Figure 4.11. This relationship highlights that metabolic pathways converge at acetyl-CoA, a common metabolic intermediate. Carbohydrates, fats, and most proteins used for energy are converted to acetyl-CoA, which then enters the TCA cycle for complete oxidation.
Figure 4.11 The relationship between CHO, FAT, and PROTEIN metabolism. The common metabolic intermediate is acetyl CoA.
When protein is combusted in a laboratory setting, it yields 5.65 kcal/g. However, within the body, energy must be expended to convert nitrogen to urea during protein metabolism, resulting in an energy yield of approximately 4.1 kcal/g[16].
Summary of Substrate Metabolism
The ability to produce muscle contraction depends on ATP synthesis, which is generated through the metabolism of carbohydrates, fats, and, in some cases, proteins. These substrates are broken down through catabolism in three energy systems: the CrP-ATP system, glycolysis, and mitochondrial respiration (oxidative phosphorylation). Carbohydrate metabolism, highlighted in this chapter, is discussed in four stages:
Glycolysis
Conversion of Pyruvate to Acetyl-CoA
TCA Cycle
Electron Transport Chain (ETC)
The CrP-ATP system and glycolysis occur in the cytosol, while oxidative phosphorylation takes place within the mitochondria. Under aerobic conditions, carbohydrates and fats are reduced to the common intermediate acetyl-CoA, which then enters the TCA cycle for complete oxidation. The hydrogens removed during redox reactions are utilized by the ETC, where the majority of ATP is generated.
Table 4.5 The factors known to affect rate-limiting enzyme activity. The common stimulator of metabolism is ADP. The common inhibitor of metabolism is ATP.
Pathway
Rate-limiting enzyme
Stimulators
Inhibitors
Phosphogen
Creatine Kinase
ADP
ATP
Glycolysis
PFK
AMP, ADP, Pi, decrease in pH
ATP, Creatine Phosphate, Citrate, increase in pH
TCA Cycle
Isocitrate dehydrogenase
ADP, Ca2+, NAD+, ADP, Pi
ATP, NADH+H+
Electron Transport Chain
Cytochrome oxidase
ADP, Pi
ATP
Chapter Summary
In this chapter, we explored the fundamental processes of energy metabolism essential for muscle contraction and overall cellular function. We began by examining the three primary energy systems: the CrP-ATP system, glycolysis, and mitochondrial respiration (oxidative phosphorylation). Each system plays a crucial role in ATP production, with specific pathways and regulatory mechanisms. Carbohydrate metabolism was also discussed, detailing its four stages: glycolysis, conversion of pyruvate to acetyl-CoA, the TCA cycle, and the electron transport chain (ETC). We highlighted the differences between glycolysis and glycogenolysis, emphasizing their ATP yields and regulatory enzymes. The chapter also covered the phosphagen system’s role in rapid ATP production and its regulation by cellular constituents.
The electron transport chain was discussed in depth, illustrating its importance in oxidative phosphorylation and ATP synthesis. We also explored beta oxidation, explaining how fatty acids are metabolized to produce ATP, and the significant energy yield from fat oxidation compared to carbohydrates and proteins. Finally, we touched on protein metabolism, noting its contribution to ATP production during prolonged activity and the conditions under which it becomes significant. The chapter concluded by analyzing the interconnectedness of carbohydrate, fat, and protein metabolism, with acetyl-CoA serving as a common metabolic intermediate.
Overall, this chapter provided a comprehensive understanding of the biochemical pathways involved in energy metabolism, highlighting their relevance to exercise physiology and cellular energy production.
Scholarly Questions
Identify the rate-limiting enzymes for the CrP-ATP system, glycolysis, the TCA cycle, and the ETC.
List the three energy systems that supply ATP.
Which energy system is the most immediate? Which system is utilized for long-duration exercise?
How long can ATP be stored in the body?
What is the chemical formula for glucose?
Define the following terms: glycogen, glucose, metabolism, glycogenolysis, gluconeogenesis, metabolic pathway, glycolysis, creatine kinase, ATPase, and PFK.
Describe the process where a phosphate is cleaved from the ATP molecule with water. What enzyme catalyzes ATP hydrolysis?
Which energy system is used for long-term energy? What metabolic pathways occur in the mitochondria?
What is the common metabolic intermediate?
What is the preferred substrate (carbohydrate, fat, or protein) for energy in the body, and why?
What is the total ATP yield from the metabolism of one glucose molecule? What about glycogen?
How many steps are there in glycolysis? In which part of the cell does glycolysis occur?
What is a hydrogen atom composed of?
Define oxidation and reduction.
Explain the function of the coenzyme carriers NAD+ and FAD. Is there a difference between NAD+ function in the cytosol and the mitochondria?
How many net ATP are produced in glycolysis?
Which energy system can produce ATP both aerobically and anaerobically?
What are the four stages of carbohydrate metabolism?
How many pyruvate molecules are produced from one glucose molecule?
Draw and label the structure of a mitochondrion. What metabolic processes occur inside the mitochondrion?
Name the protein complexes in the ETC. How many are there?
Diagram all four stages of carbohydrate metabolism and write out the ETC in a complete essay. Tally the total ATP production from one glucose molecule.
What is beta oxidation? How many ATP are produced from a 16-carbon fatty acid? An 18-carbon fatty acid?
Can protein be used to create ATP? What must protein be converted to in order to create ATP (hint: common intermediate and ?).
Robergs RA, Ghiasvand F, Parker D, The biochemistry of exercise-induced metabolic acidosis. Am J Physiol Regul Integr Comp Physiol, 2004. 287: p. R502-R516. ↵
Robergs RA, Ghiasvand F, Parker D, The biochemistry of exercise-induced metabolic acidosis. Am J Physiol Regul Integr Comp Physiol, 2004. 287: p. R502-R516. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Powers SK, and Howley ET, Exercise Physiology (Theory and Application to Fitness and Performance). 9th Edition ed. 2015, New York, NY: McGraw-Hill. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Haff GG, Triplett NT, ed., Essentials of strength training and conditioning, 4th edition/National Strength and Conditioning Association. 2016, Human Kinetics: Champaign, IL. ↵
Haff GG, Triplett NT, ed., Essentials of strength training and conditioning, 4th edition/National Strength and Conditioning Association. 2016, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Brooks G, Amino acid and protein metabolism during exercise and recovery. Med Sci Sports Exerc, 1987. 19: p. S150-S156. ↵
Graham TE, Rush JWE, MacLean DA, Skeletal muscle oxidative enzyme enhancement with endurance training. Exercise Metabolism, ed. S.L. Hargreaves M. 2006, Champaign, IL: Human Kinetics. 41-72. ↵
Haff GG, Triplett NT, ed., Essentials of strength training and conditioning, 4th edition/National Strength and Conditioning Association. 2016, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
definition
The primary energy carrier in all living organisms. It is a nucleotide composed of: adenine (a nitrogenous base), ribose (a five-carbon sugar), and three phosphate groups linked by high-energy bonds.
(Also called the ATP–PCr system) is the body’s fastest energy system, providing immediate energy for short-duration, high-intensity activities (e.g., sprinting, heavy lifting).
A metabolic pathway that breaks down glucose (C₆H₁₂O₆) into two molecules of pyruvate, producing a small amount of energy in the form of ATP and NADH.
The series of biochemical processes that occur within the mitochondria of eukaryotic cells to produce energy in the form of ATP (adenosine triphosphate).
A subatomic particle found in the nucleus of an atom. It carries a positive electric charge of +1 elementary charge and has a mass of approximately 1 atomic mass unit (1.67 × 10⁻²⁷ kg).
A nucleotide composed of three components: adenine (a nitrogenous base), ribose (a five-carbon sugar), and one phosphate group.
A simple sugar (monosaccharide) with the molecular formula C₆H₁₂O₆.
The biochemical process of breaking down glycogen into glucose molecules to provide energy for cellular metabolism. It primarily occurs in the liver (to maintain blood glucose levels) and skeletal muscle (to supply energy during muscle activity).
Membrane-bound organelles found in most eukaryotic cells, often referred to as the "powerhouses of the cell" because they produce the majority of the cell’s usable energy in the form of ATP (adenosine triphosphate).
A critical metabolic step that links glycolysis to the TCA cycle (citric acid cycle; Krebs cycle) in aerobic respiration. This process occurs in the mitochondrial matrix.
Also known as the citric acid cycle or Krebs cycle, is a central metabolic pathway that takes place in the mitochondrial matrix of eukaryotic cells. It oxidizes acetyl-CoA (derived from carbohydrates, fats, and proteins) into carbon dioxide (CO₂) while generating high-energy electron carriers and a small amount of ATP.
A series of protein complexes and associated molecules located in the inner mitochondrial membrane that drive oxidative phosphorylation, the final stage of aerobic respiration.
The catabolic process by which fatty acids are broken down in the mitochondria (or peroxisomes) to generate acetyl-CoA, which then enters the TCA cycle for energy production.
The metabolic pathway that synthesizes glucose from non-carbohydrate precursors, ensuring a continuous supply of glucose during fasting, prolonged exercise, or low-carbohydrate intake. It primarily occurs in the liver (and to a lesser extent in the kidney cortex).