The major components of the respiratory system and a magnified view of the alveoli and its associated capillary beds.
Learning Objectives
Explain the interdependence of the pulmonary and cardiovascular systems.
Identify the four processes involved in the transportation of oxygen and carbon dioxide in the body.
Define pulmonary ventilation and pulmonary diffusion and the processes of external and internal respiration.
Identify the major organs of the respiratory system and their locations.
Outline the pathway of environmental air from the nose to the alveoli.
Explain the roles of the conducting and respiratory zones.
Explain the principles of Boyle’s Law and Fick’s Law of diffusion as they apply to respiration.
Identify Factors Affecting Oxygen and Carbon Dioxide Transport.
Explain the Bohr and Haldane effects.
Define and measure tidal volume, vital capacity, residual volume, and total lung capacity.
Describe the changes in ventilation during incremental exercise and the concept of the ventilatory threshold.
Explain the effects of exercise in hot environments on ventilation.
Discuss the neural and chemoreceptor control of ventilation during and after exercise.
Introduction
The pulmonary and cardiovascular systems work in tandem to form an efficient delivery mechanism that transports oxygen to tissues and removes carbon dioxide. Due to their close interdependence, these systems are often collectively referred to as the cardiopulmonary system. The transportation of oxygen and carbon dioxide within the body encompasses four distinct processes:
Pulmonary Ventilation
Pulmonary Diffusion
Transportation via the Blood
Capillary Diffusion
Pulmonary ventilation, commonly known as breathing, involves the movement of air into and out of the lungs. During pulmonary diffusion, the exchange of oxygen and carbon dioxide occurs between the lungs and the blood. This process replenishes the blood’s oxygen supply, which is depleted at the tissue level for oxidative energy production, and removes carbon dioxide from the systemic venous blood. In the lungs, oxygen is loaded into the blood while carbon dioxide is unloaded and exhaled. Both pulmonary ventilation and pulmonary diffusion are forms of external respiration, which refers to gas exchange occurring in the alveoli and capillary beds of the lungs[1]. Following pulmonary diffusion, gases are transported by the blood to various tissues. Upon reaching the tissues, gases diffuse through the capillaries into the tissues. This gas exchange process at the tissue level is known as internal respiration or cellular respiration. The circulatory system links both internal and external respiration, highlighting the integral role of the pulmonary system in gas exchange.
Understanding lung function is crucial due to its significant role in this process. This chapter will explore the design and components of respiration and describe the ventilatory responses to exercise.
Anatomy of the Respiratory System
Figure 10.1 The major organs of the respiratory system.
The lungs consist of a network of respiratory passages that facilitate the close association of lung capillaries with environmental air. This proximity enables the efficient exchange of oxygen and carbon dioxide with the atmosphere. The primary organs of the respiratory system are illustrated in Figure 10.1.
Located in the thoracic cavity, the lungs flank the heart laterally and are the main organs of the respiratory system. They are not directly attached to the ribs but are suspended by connective tissue sacs known as pleural sacs. The pleura is a slick, sticky, semi-transparent serous membrane that provides lubrication for the movable lungs and heart (Figure 10.2). The pleurae produce a watery fluid called pleural fluid. The pleurae serve three main purposes:
Reducing Friction: Since the heart, lungs, and diaphragm are in constant motion, the pleurae minimize friction between the ribs and these organs.
Creating a Pressure Gradient: The pressure within the pleurae is lower than atmospheric pressure, creating a pressure gradient essential for gas exchange (to be discussed later in the chapter).
Compartmentalization: The pleurae separate the abdominal cavity from the thoracic cavity, compartmentalizing the heart and lungs and preventing infections from spreading from other organs to the thoracic cavity.
When the pleurae do not function correctly, it can result in painful and difficult breathing.
Figure 10.2 The anatomy of the lungs, the pleural sack.
Figure 10.2 illustrates the three components of the pleurae. The most superficial layer is the parietal pleurae, which covers the diaphragm and attaches to the internal walls of the thoracic cavity. Beneath the parietal pleurae lies the pleural cavity, containing serous fluid. Each thorax holds approximately 16 ml of serous fluid, which drains from the chest via the lymphatic vessels. This fluid lubricates the moving parts within the chest. The deepest layer is the visceral pleurae, which attaches directly to the lungs. Together, the pleurae connect the surfaces of the lungs and ribs, preventing lung collapse when the ribs expand.
The lung tissue, or spongy parenchyma, contains the bronchial tree and allows for the elastic recoil of the lungs after expansion during inspiration. This tissue includes a significant proportion of elastic fibers, which aid in this recoil.
The Pathway of Environmental Air into the Body
Environmental air enters the body through the nose and nasal cavity. As the air travels through the body, it is warmed, cleansed, and humidified by swirling through the irregular sinus surfaces, causing dust and other particles to adhere to the nasal mucosa. This process filters out most particles that could infect the respiratory tract. From the nasal cavity, air moves down a series of rigid pathways commonly known as the “windpipe,” which includes the pharynx, larynx, and trachea. These structures are lined with cartilage, providing an open airway for the passage of gases.
Figure 10.3 The conducting zone is composed of the trachea, bronchial tree, and bronchioles.
Air travels from the trachea into the left and right lungs, entering the bronchial tree. The bronchial tree consists of progressively smaller airways, culminating in approximately 65,000 terminal bronchioles. The primary bronchus is the largest airway, which then branches into secondary bronchioles. These secondary bronchioles further divide into tertiary bronchi and smaller bronchioles. These structures form the conducting zone of the lungs, responsible for directing air towards the respiratory zone (Figure 10.3). It is important to note that gas exchange does not occur in the conducting zone; its primary function is to maintain an open pathway for air to reach the respiratory zone.
From the bronchioles, air moves into the terminal bronchioles (also known as respiratory bronchioles), which have scattered alveoli in their walls. Unlike the windpipe, terminal bronchioles are lined with smooth muscle, allowing the airways to dilate or constrict. The ends of the terminal bronchioles consist of “sacs” of alveoli that line the alveolar ducts. These alveolar sacs are surrounded by capillary networks, facilitating the oxygenation of blood and the release of carbon dioxide into the lungs as waste. Together, the terminal bronchioles and alveolar sacs constitute the respiratory zone (Figure 10.4). The respiratory zone is where gas exchange between air and blood occurs, and where external respiration takes place.
Gas exchange in the lungs occurs across approximately 300 million alveoli. Due to their vast number, these tiny alveoli provide the lungs with a large surface area for diffusion. Each alveolus is only one cell thick, which enhances the diffusion of gases at the respiratory membrane. According to Fick’s law of diffusion, the rate of gas diffusion across tissues is inversely proportional to the tissue thickness.
Figure 10.4 The respiratory zone is comprised of the terminal bronchioles and the alveolar sacs.
Alveoli contain specialized cells that facilitate their function. The cells lining the alveoli are known as simple squamous cells, which also form the respiratory membrane between the pulmonary capillaries. The respiratory membrane, measuring 0.5 micrometers in diameter, consists of two basement membranes sandwiched between a simple squamous cell lining the alveolus and a simple squamous cell lining the capillary wall. This membrane is only two cells thick, making it very fragile and susceptible to damage from smoking, pollution, and disease.
Figure 10.5 The enlarged view of an alveolus showing its components and the regions of gas exchange between the alveolus and the pulmonary blood in the capillaries.
Alveolar macrophages, also known as “dust cells,” play a crucial role in eliminating bacteria, dust, pollen, and pollutants that reach the respiratory zone. The air in the alveoli is humidified before arrival, which creates a challenge for the lungs. The surface tension of the liquid in the air is relatively high, posing a risk of alveolar collapse.
To counteract this, type II alveolar cells produce a substance called pulmonary surfactant. Surfactant reduces the surface tension of water in the alveoli, preventing their collapse due to the attractive forces of water[2]. Figure 10.5 illustrates the relationship between the different types of cells that make up an alveolus.
The Respiratory Muscles
Respiratory muscles are skeletal muscles that act upon the chest wall to facilitate the movement of gas in and out of the lungs. During resting ventilation, the diaphragm and external intercostal muscles drive the volume changes in the thoracic cavity essential for breathing.
The external intercostal muscles are the most superficial layer of muscles located between the ribs. When they contract, they assist in lifting the rib cage away from the abdomen. Unfortunately, spinal cord injuries that damage the phrenic nerve can result in the inability to operate the diaphragm, leading to an inability to ventilate.
The diaphragm is the most important muscle for ventilation and is the only skeletal muscle considered essential for life. It is a concave-shaped muscle located beneath the lungs and attached to the ribs (Figure 10.6). When the diaphragm contracts, it moves downward, forcing the abdominal contents downward and forward.
Figure 10.6 The inferior view of the diaphragm and associated anatomical features.
During resting ventilation, the external intercostal muscles and the diaphragm are the primary muscles involved in breathing. However, during exercise, ventilation increases to meet the muscles’ oxygen demands and to expel carbon dioxide. This results in an increase in pulmonary ventilation, placing a greater workload on the respiratory muscles.
During exercise and deep breathing, accessory muscles also contribute to the intake and expulsion of air. The muscles involved in deep inspiration include the scalene muscles, pectoralis minor, and the sternocleidomastoid, in addition to the diaphragm and external intercostal muscles [2]. These muscles assist the diaphragm in increasing the volume of the chest, thereby aiding in inspiration (see pulmonary ventilation).
Figure 10.7 The muscles of deep inspiration.
Typically, expiration is a passive process that does not require ATP. However, during deep expiration and expiration associated with exercise, accessory muscles are engaged, making it an active process. The muscles involved in deep expiration are those of the abdominal wall, including the internal intercostal muscles, external oblique, internal oblique, and transverse abdominis (Figure 10.8). When these muscles contract, they push the diaphragm upward and pull the ribs downward and inward. This action decreases the dimensions of the thoracic cavity, aiding in the expulsion of air.
Figure 10.8 The muscles of deep expiration.
Ventilation Physiology
The movement of air from the environment into the lungs is known as pulmonary ventilation and occurs due to changes in pressure and volume. Resting inhalation is an active process that requires ATP, as it involves the contraction of the diaphragm and external intercostal muscles. Conversely, resting exhalation is a passive process that does not require ATP, as it involves the relaxation of these muscles.
To understand the mechanics of ventilation, it is essential to grasp Boyle’s Law. Boyle’s Law states that the absolute pressure exerted by a given mass of an ideal gas is inversely proportional to the volume it occupies, provided the temperature and the amount of gas remain constant within a closed system (i.e., ( P1V1 = P2V2 ))[3].
Figure 10.9 illustrates Boyle’s Law using containers of specific volumes and a precise number of particles. In a larger container with a fixed number of particles, the pressure exerted by the particles is relatively low due to their interactions with each other and the container. When the volume of the container decreases while maintaining the same number of particles, the pressure exerted by the particles on themselves and the container increases. Thus, pressure and volume are inversely related.
Figure 10.9 Containers of particles that demonstrate Boyle’s law by showing the inverse relationship between pressure and volume.
Simple diffusion refers to the process by which gases, such as carbon dioxide and oxygen, pass through a membrane without the assistance of intermediary molecules like integral membrane proteins. The driving force behind this movement is the force of diffusion. In terms of gas pressures, this means that gases will diffuse from an area of higher pressure to an area of lower pressure until the pressures on both sides of the membrane are equal, a state known as equilibrium.
Therefore, based on the principles of simple diffusion, a pressure gradient (i.e., the difference in pressures across a membrane) is essential for gas exchange to occur.
Figure 10.10 The physiological mechanisms of inspiration and expiration.
By applying the principles of Boyle’s Law and simple diffusion to the lungs, the mechanisms of ventilation can be understood (Figure 10.10). During resting inhalation, the diaphragm contracts and moves downward, enlarging the volume of the thoracic cavity. The external intercostal muscles also contribute to this enlargement. This increase in cavity volume reduces intrapulmonary pressure below the environmental air pressure, creating a suction force that draws air into the lungs to equalize the pressure difference.
Expiration occurs in the opposite manner. During expiration, the diaphragm and external intercostal muscles relax, decreasing the volume of the thoracic cavity and increasing intrapulmonary pressure. This makes the pressure inside the lungs greater than the environmental pressure, resulting in the expulsion of air from the lungs.
During forced or labored breathing, such as during heavy exercise, inspiration and expiration are assisted by accessory muscles (see respiratory muscles). The pressure changes required for ventilation at rest are relatively small. The standard atmospheric pressure at sea level is 760 mmHg. Inspiration may decrease the pressure in the lungs by 2 to 3 mmHg. However, during exhaustive exercise, intrapulmonary pressure can decrease by up to 100 mmHg[4].
Partial Pressures of Physiological Gases
Before discussing the diffusion of gases across membranes in the body, it is essential to understand the concept of partial pressures. Atmospheric air is a mixture of oxygen, carbon dioxide, nitrogen, and trace amounts of other gases. Each gas exerts its own pressure, known as the partial pressure (P). According to Dalton’s Law, the total pressure of a mixture of gases is equal to the sum of the partial pressures of the individual gases.
This concept is illustrated in Figure 10.11, where the total pressure of two gases in the same container can be calculated by adding their partial pressures. For example, if the partial pressure of oxygen is 159 mmHg and the partial pressure of nitrogen is 593 mmHg, the total pressure of the gas mixture is 752 mmHg (i.e., 593 mmHg + 159 mmHg = 752 mmHg).
Figure 10.11 Three containers of oxygen, nitrogen, and mixed gas particles and their additive pressures demonstrating partial pressures, total pressure, and Dalton’s law.
The partial pressure of a gas is proportional to its concentration. Atmospheric air is generally composed of 20.93% oxygen, 79.04% nitrogen, and 0.03% carbon dioxide, totaling 100%. To determine the partial pressures of each gas, it is necessary to know the barometric pressure (PB), which is the force exerted by the weight of the gas in the atmosphere. At sea level, the barometric pressure is typically 760 mmHg.
For example, to calculate the partial pressure of oxygen (PO2) in air at sea level, first determine the fraction of air composed of oxygen. Since oxygen makes up 20.93% of the air, the fraction is 0.2093 (20.93/100). The partial pressure of oxygen at sea level can then be computed as:
PO2 = 760 mmHg x 0.2093
PO2 = 159 mmHg
Fick’s Law of Diffusion
Another important concept in the diffusion of gases in the body is Fick’s Law of Diffusion. Fick’s Law of Diffusion states that the rate of gas transfer (V) is proportional to the tissue area (A), the diffusion coefficient of the gas (D), and the difference in the partial pressure of the gas on the two sides of the tissue (P1 – P2), and is inversely proportional to the membrane thickness (T):
Vgas = (A/T) x D x (P1 – P2)
Therefore, a pressure gradient is necessary to move gases from one part of the body to another. A pressure gradient is the difference in partial pressures on opposite sides of a membrane. The lung is well-designed for the diffusion of gases across the alveolar membrane, with an estimated surface area of 60 to 80 square meters (approximately the size of a tennis court). Additionally, the alveolar membrane is extremely thin. Together, these features make the lung an ideal organ for gas exchange. This is crucial because gas exchange throughout the body depends on the factors of diffusion, primarily driven by pressure gradients.
Figure 10.12 Partial pressure changes of the atmosphere air, alveolar air, pulmonary circuit, and systemic circuit.
Figure 10.12 illustrates gas exchange via diffusion across the alveolar-capillary membrane and at the capillary-tissue membranes. The partial pressure of carbon dioxide (PCO2) and the partial pressure of oxygen (PO2) in the alveolar capillaries are 40 mmHg and 104 mmHg, respectively. In contrast, the PCO2 in the atmosphere is 0.2 mmHg, and the PO2 is 159 mmHg. This creates a PO2 difference of 55 mmHg in the alveoli and a PCO2 difference of 39.8 mmHg in the atmosphere. Consequently, due to the pressure gradient, carbon dioxide leaves the alveoli and diffuses into the alveolus, while oxygen diffuses into the alveoli.
Oxygen Transport
Hemoglobin, a protein in red blood cells, transports oxygen. When bound to oxygen, it is called oxyhemoglobin; when not bound to oxygen, it is referred to as deoxyhemoglobin. The loading and unloading of hemoglobin are reversible reactions:
Deoxyhemoglobin + O2 ⇔ Oxyhemoglobin
Approximately 98% of oxygen in the blood is transported as oxyhemoglobin, while the remaining 2% is dissolved in the plasma, as oxygen is a poor solute. Oxygen unloading from hemoglobin is determined by the partial pressures of the tissues and other factors.
Each hemoglobin molecule can bind four oxygen molecules, and the amount of oxygen transported per volume of blood depends on the concentration of hemoglobin. In healthy males, the concentration of hemoglobin is approximately 150 grams per liter of blood, and in females, it is about 130 grams per liter[5]. When fully saturated, each gram of hemoglobin can transport 1.34 ml of oxygen. Therefore, at 100% saturation, a healthy adult can transport between 174-200 ml of oxygen per liter of blood.
The oxygen-hemoglobin (O2-Hb) dissociation curve, shown in Figure 10.13, demonstrates the relationship between oxygen and hemoglobin binding. The O2-Hb dissociation curve is sigmoidal (S-shaped) and describes the amount of oxygen unloaded at the tissues. The percent hemoglobin saturated with oxygen increases sharply up to an arterial PO2 of 40 mmHg. At PO2 values above 40 mmHg, hemoglobin saturation rises slowly to a plateau around 90-100 mmHg, where hemoglobin is approximately 98% saturated.
At rest, the body’s oxygen requirements are relatively low, and only about 25% of oxygen is unloaded at the tissues, as shown in Figure 10.13 when the PO2 at the tissues is 40 mmHg. However, during strenuous exercise, the muscles’ demand for oxygen increases, which can drop the PO2 in the tissues to 20 mmHg. In such cases, the muscles and peripheral tissues can extract up to 90% of the oxygen bound to hemoglobin.
Figure 10.13 The oxyhemoglobin dissociation curve.
Several factors influence both blood PO2 and the unloading of oxygen at the tissues. Changes in acidity, increases in PO2, temperature, and levels of 2-3-diphosphoglyceric acid (2,3-DPG) all enhance oxygen unloading (i.e., oxygen dissociation from hemoglobin) at the tissues. This is particularly important during exercise, as efficient oxygen unloading is crucial for the continued production of ATP.
The bond strength between oxygen and hemoglobin is weakened by a decrease in blood pH, leading to increased oxygen unloading to the tissues. This phenomenon is represented by a “rightward” shift in the oxyhemoglobin dissociation curve and is known as the Bohr effect (Figure 10.14). During heavy exercise, the rise in blood hydrogen ion levels contributes to this effect. Physiologically, hydrogen ions in the blood compete for binding sites on hemoglobin, reducing its oxygen transport capacity. High concentrations of hydrogen ions (e.g., during acidosis) cause a reduction in hemoglobin’s affinity for oxygen.
An increase in core temperature also causes a rightward shift in the curve, as this condition weakens the bond between oxygen and hemoglobin. Conversely, a decrease in blood temperature results in a stronger bond, hindering oxygen release at the tissues. As shown by the resting curve (green line) in Figure 10.14, the curve can shift to the left or right depending on these factors.
Figure 10.14 The Bohr effect and factors that affect the oxygen hemoglobin disassociation curve.
Carbon dioxide in the blood can displace oxygen from hemoglobin by binding with the amine radicals of the hemoglobin molecule. This increase in blood PCO2 contributes to the Bohr effect, which enhances oxygen unloading at the tissues. Conversely, the binding of oxygen to hemoglobin tends to displace carbon dioxide from the blood, known as the Haldane effect. This occurs because the combination of oxygen with hemoglobin in the lungs makes hemoglobin a stronger acid, which displaces carbon dioxide from the blood. As a result, the increased acidity of hemoglobin causes excess hydrogen ions transported by red blood cells to be released in the lungs. These hydrogen ions bind with bicarbonate to form carbonic acid, which then dissociates into water and carbon dioxide. The carbon dioxide is subsequently released from the blood into the alveoli and finally exhaled[6]. Thus, oxygen loading in the lungs facilitates carbon dioxide unloading.
Another factor affecting the hemoglobin dissociation curve is 2,3-diphosphoglycerate (2,3-DPG), a byproduct of glycolysis. Red blood cells rely exclusively on glycolysis for energy production, leading to the creation of 2,3-DPG when they are metabolically active. 2,3-DPG binds with hemoglobin and reduces its affinity for oxygen. The production of 2,3-DPG increases with exposure to anemia and high altitudes, due to the reliance on glycolysis under lower PO2 conditions.
When oxygen is delivered to muscles, myoglobin, another oxygen-binding protein, shuttles oxygen from the muscle cell membrane to the mitochondria. Myoglobin is one-fourth the weight of hemoglobin and has a higher affinity for oxygen, even at lower PO2. Myoglobin releases its oxygen only at very low PO2, compatible with the PO2 of exercising muscle, which can be as low as 1 to 2 mmHg[7]. Type I muscle fibers have a higher concentration of myoglobin than Type II fibers. Myoglobin may serve as an “O2 reservoir” for muscles during the transition from rest to exercise. It is thought that at the end of exercise, myoglobin oxygen stores are replenished, contributing to the oxygen debt (EPOC).
Carbon Dioxide Transport
Carbon dioxide, produced by tissues during cellular respiration, diffuses out of cells in its dissolved molecular form. Upon entering the tissue capillaries, carbon dioxide undergoes instantaneous physical and chemical reactions for transport. Carbon dioxide is transported in the blood in three forms:
Approximately 10% is dissolved in the plasma.
20% is bound to hemoglobin (forming carbaminohemoglobin).
70% is transported as bicarbonate ion (HCO3-).
Most of these processes occur within red blood cells (Figure 10.15). Carbon dioxide can be transported in far greater quantities than oxygen, significantly influencing the acid-base balance of body fluids. Under resting conditions, an average of 4 ml of carbon dioxide is transported from the tissues to the lungs per 100 ml of blood[8]. The majority of carbon dioxide produced by active muscles is transported back to the lungs as bicarbonate ion.
A high PCO2 in the blood causes carbon dioxide to combine with water to form carbonic acid. This reaction would occur too slowly without the enzyme carbonic anhydrase inside red blood cells, which catalyzes the reaction and accelerates its rate approximately 5,000 times. Carbonic acid (H2CO3) quickly dissociates into bicarbonate ion (HCO3-) and a hydrogen ion (H+). The hydrogen ion binds to hemoglobin in red blood cells, as hemoglobin is a powerful acid-base buffer. The bicarbonate ion then diffuses into the plasma, where it is transported. Chloride ions diffuse into red cells to replace the bicarbonate, a phenomenon known as the chloride shift. This results in a higher chloride content in venous red blood cells compared to arterial red cells.
Figure 10.15 Summary of carbon dioxide transport in the blood from the tissues.
In addition to reacting with water, some carbon dioxide reacts directly with the hemoglobin molecule to form carbaminohemoglobin (CO2Hgb). This reaction is reversible, allowing carbon dioxide to be easily released into the alveoli where PCO2 is lower than in the pulmonary capillaries. However, this reaction is much slower than the reaction between carbon dioxide and water, so under normal conditions, the carbamino-mechanism of transport accounts for no more than 20% of the total carbon dioxide transported[9].
At the lungs, the PCO2 of the blood is greater than that in the alveolus, causing carbon dioxide to diffuse out of the blood across the membrane. When the blood reaches the pulmonary capillaries, carbonic acid dissociates into carbon dioxide and water, with the carbon dioxide being exhaled.
Pulmonary Volumes and Capacities
The volume of air in the lungs can be measured using a technique called spirometry. In this method, the subject breathes into a device capable of measuring both inspired and expired gas volumes. Spirometry assesses the rate of expired airflow and lung capacity, making it a valuable tool for diagnosing lung diseases such as chronic obstructive pulmonary disease (COPD) and asthma.
A modern spirometer, shown in Figure 10.16, includes a mouthpiece similar to the one used in indirect calorimetry. Several measures of pulmonary function can be obtained through spirometry, including the total lung capacity and tidal volume of an individual.
Figure 10.16 Device into which a patient breathes for Spirometry, Body Plethysmography and other related medical tests. Photographed at Swedish Hospital Ballard Campus, Seattle, Washington.
The volume of air that enters and leaves the lungs during normal, resting conditions with each breath is called the tidal volume (VT) . Tidal volume typically displaces about 500 ml of air between inhalation and exhalation. Pulmonary ventilation (VE) refers to the volume of air moved in or out of the lungs per minute. It is the product of tidal volume (VT) and breathing frequency (f). Mathematically, ventilation can be expressed as:
VE = VT x f
This equation helps quantify the amount of air exchanged in the lungs over a given period, providing valuable insights into respiratory function.
Figure 10.17 graphically illustrates the measurement of tidal volumes during normal quiet breathing and the various lung volumes and capacities. The vital capacity (VC) is the greatest amount of air that can be expired following a maximal inspiration, typically about 4,700 ml. Even after a maximal voluntary expiration, a small volume of air remains in the lungs, known as the residual volume (RV), which is about 1,300 ml and cannot be voluntarily exhaled. The sum of the residual volume and the vital capacity constitutes the total lung capacity (TLC), which is approximately 6 liters in healthy individuals:
TLC = RV + VC
Figure 10.17 Lung volumes at rest measured by spirometry (a). Inspiratory capacity, functional capacity vital capacity, residual volume, and total lung capacity are shown (b).
Pulmonary Ventilation During Dynamic Exercise
The onset of exercise is accompanied by an increase in ventilation. Initially, ventilation increases rapidly and then rises more slowly towards a steady state, even without an initial increase in arterial PCO2. The initial rise in ventilation is controlled by the nervous system and mediated by respiratory control centers in the brain. The more gradual rise towards steady-state exercise is controlled by changes in the chemical status of the arterial blood. Increased metabolism in skeletal muscles raises blood PCO2. Increases in PCO2 and hydrogen ions are sensed by chemoreceptors in the brain, carotid bodies, and lungs, which stimulate the inspiratory center to increase the frequency and depth of respiration. Similar to heart rate, there may also be an anticipatory effect where ventilation rises before exercise commences.
During strenuous exercise, oxygen consumption and carbon dioxide formation can increase up to 20-fold[10]. Maximal ventilation rates of approximately 100 L/min are common for smaller individuals but may exceed 200 L/min in larger individuals [1]. During mild, steady-state exercise, ventilation increases to match the rate of energy expenditure. The ratio of air ventilated to oxygen consumed in a given time is the ventilatory equivalent for oxygen (VE/VO2). At rest, this ratio ranges from 23-28 L of air per liter of oxygen. This value changes very little during low-intensity exercise, such as walking. However, at near-maximal exercise intensities, the ventilatory equivalent for oxygen can exceed 30 L of air per liter of oxygen. Generally, the ventilatory equivalent for oxygen remains relatively constant, indicating that the control of breathing is properly matched to the muscles’ demand for oxygen.
Where (Q) is blood flow, or perfusion. The ideal ventilation-perfusion ratio is 1.0, indicating a perfect match between ventilation and cardiac output. However, gas exchange is not always perfect in the exercising lung[11]. Light-to-moderate exercise intensities improve ventilation-perfusion matching. During heavy exercise, small ventilation-perfusion inequalities may occur because red blood cells do not have enough transit time in the lung capillaries due to high heart rates. This can impair gas exchange and cause a condition known as exercise-induced hypoxemia.
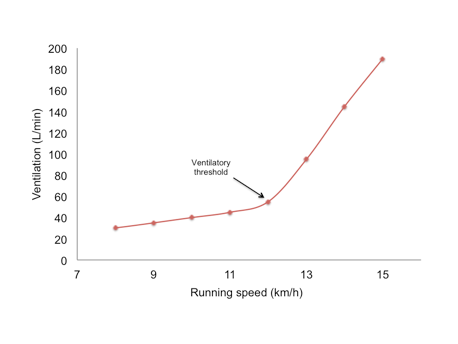
Figure 10.18 Changes in pulmonary ventilation during running at increasing speeds, illustrating the concept of the ventilatory threshold. Ventilatory threshold (Tvent) is the inflection point where ventilation increases exponentially.
During incremental exercise with increasing speeds, ventilation increases linearly up to 50-70% of VO2max. Beyond this point, there is a disproportionate increase in ventilation, known as the ventilatory threshold, illustrated in Figure 10.18. The ventilatory threshold reflects the respiratory response to increased carbon dioxide levels, causing a dramatic rise in ventilation. In trained runners, the ventilatory threshold occurs at a higher work rate. Trained individuals can also maintain their pH and have lower lactate levels at higher work rates compared to untrained individuals[12].
Ventilation in Hot Environments
Exercise in a hot environment makes it challenging to maintain steady-state ventilation. During prolonged sub-maximal exercise in the heat, ventilation may drift upward despite little to no change in PO2. This increase is not due to elevated PCO2 but is likely caused by catecholamines and increased core temperature. This phenomenon is known as VO2 drift.
Neural and Chemoreceptor Control of Ventilation
Input to the respiratory control center to increase ventilation can come from both neural and chemoreceptor sources. Central chemoreceptors in the medulla sense changes in PCO2 and hydrogen ion concentration. Peripheral chemoreceptors located in the aorta and carotid artery detect changes in PCO2, hydrogen ion concentration, and PO2.
At the end of exercise, pulmonary ventilation returns to normal more slowly than the decrease in the muscles’ energy demands. Post-exercise breathing rate remains elevated, primarily regulated by acid-base balance, PCO2, and blood temperature.
Chapter Summary
This chapter provides a comprehensive overview of the cardiopulmonary system, emphasizing the interdependence of the pulmonary and cardiovascular systems in transporting oxygen and carbon dioxide. It outlines the four key processes involved in gas transportation: pulmonary ventilation, pulmonary diffusion, transportation via the blood, and capillary diffusion. Pulmonary ventilation, or breathing, involves the movement of air into and out of the lungs, while pulmonary diffusion refers to the exchange of gases between the lungs and blood. These processes are crucial for replenishing blood oxygen levels and removing carbon dioxide.
The lungs, located in the thoracic cavity, are suspended by pleural sacs that provide lubrication and reduce friction. The pleurae also create a pressure gradient essential for gas exchange and compartmentalize the thoracic cavity to prevent infections. Air travels from the nose through the nasal cavity, pharynx, larynx, trachea, and bronchial tree, eventually reaching the alveoli where gas exchange occurs. The conducting zone directs air to the respiratory zone, where oxygen and carbon dioxide are exchanged.
Gas exchange in the alveoli is facilitated by a large surface area and thin respiratory membrane. Boyle’s Law and Fick’s Law explain the mechanics of ventilation and the rate of gas diffusion, respectively. The Bohr and Haldane effects describe how changes in blood pH and (PCO2) levels influence oxygen and carbon dioxide transport. 2,3-DPG and myoglobin also play roles in oxygen transport, particularly during exercise.
Spirometry measures lung volumes and capacities, such as tidal volume, vital capacity, residual volume, and total lung capacity. These measurements are essential for diagnosing respiratory conditions. Ventilation increases during exercise to meet the muscles’ oxygen demands and expel carbon dioxide. The ventilatory threshold marks a disproportionate increase in ventilation, reflecting the body’s response to increased (PCO2) levels. Exercise in hot environments and the role of neural and chemoreceptor control in regulating ventilation are also discussed.
Overall, this chapter equips students with a thorough understanding of respiratory physiology, the mechanics of pulmonary function, and the factors influencing gas exchange and transport in the body.
Scholarly Questions
What is the difference between internal and external respiration?
Can you name three functions of the pleurae?
Review the pathway of environmental air into the body. How is oxygen transported in the blood once it is loaded?
How is carbon dioxide transported in the body?
What are the components of the respiratory zone and the conducting zone?
Approximately how many terminal bronchioles are there in a healthy adult?
Explain Boyle’s Law and how it helps to explain ventilation.
Discuss the mechanisms of inspiration and expiration. Which muscles are involved in resting inhalation and expiration? Which muscles are involved in deep inspiration and expiration?
Is deep expiration an active or passive process?
What is a pressure gradient? What is a partial pressure?
Discuss Dalton’s Law.
Calculate the following pressure gradient: Atmospheric PO2 = 159 mmHg; Alveolar PO2 = 104 mmHg.
What are the partial pressures of nitrogen, oxygen, and carbon dioxide at the standard barometric pressure (760 mmHg)? Calculate the partial pressures at a barometric pressure of 640 mmHg.
Based on the principles of simple diffusion, will gas concentrations flow from lower to higher, or higher to lower?
How many hemoglobin molecules are present in one red blood cell?
How many oxygen molecules can bind to each hemoglobin molecule?
If the PO2 is low, will the hemoglobin be more or less saturated?
How do scientists represent the relationship between oxygen and hemoglobin binding?
According to the oxyhemoglobin dissociation curve, if the PO2 is 40 mmHg, what is the hemoglobin saturation? How much oxygen has been unloaded at the tissues?
If the PO2 is 20 mmHg, how much hemoglobin is saturated?
What does equilibrium mean? What is the equilibrium reached at the tissues?
What is the PO2 in the alveoli, tissues, and systemic veins?
What is the PCO2 in the alveoli, tissues, and systemic veins?
What factors affect oxygen loading? What is the effect called that causes a shift in the oxygen-hemoglobin dissociation curve? Discuss how exercise helps oxygen unloading.
What is the Haldane effect?
Determine the PO2 and PCO2 as blood travels through the pulmonary and systemic circuits.
How does exercise affect ventilation and perfusion?
Kenney WL, Wilmore JH, Costil DL, ed. Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Powers SK, and Howley ET, Exercise Physiology (Theory and Application to Fitness and Performance). 9th Edition ed. 2015, New York, NY: McGraw-Hill. ↵
Levine IN, Physical Chemistry. 1978, Boston, MA: McGraw Hill. ↵
Kenney WL, Wilmore JH, Costil DL, ed. Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Levitsky M, Pulmonary Physiology. 2007, New York, NY: McGraw Hill. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Kenney WL, Wilmore JH, Costil DL, ed. Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Dempsey JA, Wagner PD, Exercise-induced arterial hypoxemia. Journal of Applied Physiology, 1999. 87(6): p. 1997-2006. ↵
Acevedo EO, Goldfarb AH, Increased training intensity effects on plasma lactate, ventilatory threshold, and endurance. Medicine & Science in Sports & Exercise, 1989. 21(5): p. 563-568. ↵
definition
The process of moving air into and out of the lungs. It involves two phases: inhalation (inspiration), where air is drawn into the lungs, and exhalation (expiration), where air is expelled from the lungs.
The process by which gases are exchanged between the alveoli in the lungs and the blood in the pulmonary capillaries.
The exchange of gases between the air in the alveoli of the lungs and the blood in the pulmonary capillaries.
The exchange of gases between the blood in systemic capillaries and the body’s tissues.
A double-layered serous membrane that surrounds each lung and lines the chest cavity.
The outer layer of the pleural membrane that lines the inner surface of the thoracic cavity, including the chest wall, diaphragm, and mediastinum
A thin space filled with pleural fluid that reduces friction during breathing and allows the lungs to expand and contract smoothly.
The inner layer of the pleural membrane that directly covers the surface of each lung. It closely adheres to the lung tissue, following its contours and fissures.
The functional tissue of the lungs that has a soft, porous, and elastic structure, primarily composed of alveoli, bronchioles, and associated capillaries.
Refers to the branching system of airways within the lungs that conducts air from the trachea to the alveoli
The portion of the respiratory system where gas exchange occurs. It includes the respiratory bronchioles, alveolar ducts, alveolar sacs, and alveoli. These structures contain thin walls and are closely associated with pulmonary capillaries, allowing oxygen to diffuse into the blood and carbon dioxide to diffuse out.
The portion of the respiratory system responsible for transporting air to the lungs but not directly involved in gas exchange. It includes the nose, nasal cavity, pharynx, larynx, trachea, bronchi, and bronchioles (up to the terminal bronchioles). These structures serve to warm, humidify, and filter incoming air, ensuring that it reaches the respiratory zone in optimal condition for gas exchange.
Tiny, balloon-like air sacs located at the ends of the bronchioles in the lungs.
A type of epithelial cell characterized by their thin, flat shape and single-layer arrangement.
The thin barrier through which gas exchange occurs between the air in the alveoli and the blood in the pulmonary capillaries
Specialized immune cells located within the alveoli of the lungs. They play a critical role in the respiratory system’s defense by engulfing and digesting airborne particles, pathogens, and cellular debris through a process called phagocytosis.
Specialized epithelial cells found in the alveoli of the lungs. Their primary function is to produce and secrete pulmonary surfactant, a lipoprotein substance that reduces surface tension within the alveoli. This surfactant prevents alveolar collapse during exhalation and helps maintain lung compliance, making breathing more efficient. In addition to surfactant production, these cells also play a role in alveolar repair and regeneration.
A lipoprotein substance that reduces surface tension within the alveoli. This surfactant prevents alveolar collapse during exhalation and helps maintain lung compliance, making breathing more efficient.
A dome-shaped sheet of skeletal muscle that separates the thoracic cavity from the abdominal cavity. It plays a crucial role in respiration
A group of skeletal muscles located between the ribs that play a key role in the mechanics of breathing, particularly during inhalation
A group of three paired muscles (anterior, middle, and posterior) located in the lateral neck. During forced inhalation, they assist in elevating the first and second ribs, helping to expand the thoracic cavity and facilitate airflow into the lungs.
A thin, triangular muscle located beneath the pectoralis major in the upper chest. It attaches from the ribs (usually the 3rd to 5th) to the coracoid process of the scapula.
A prominent neck muscle that extends from the sternum and clavicle to the mastoid process of the skull. In addition to its role in head rotation and flexion, it assists in elevating the sternum during forced inhalation, thereby increasing the volume of the thoracic cavity.
Located between the ribs, deep to the external intercostals. They assist in forced exhalation by pulling the ribs downward and inward, which decreases the volume of the thoracic cavity and helps push air out of the lungs. These muscles are especially active during vigorous breathing or physical exertion.
A broad, flat muscle located on the lateral and anterior parts of the abdomen. It contributes to forced exhalation by compressing the abdominal cavity, which pushes the diaphragm upward and helps expel air from the lungs.
A muscle that lies just beneath the external oblique and runs in the opposite direction. Like the external oblique, it aids in forced exhalation by compressing the abdominal contents and assisting in diaphragm elevation.
The deepest of the abdominal muscles, wrapping horizontally around the abdomen. It plays a key role in forced exhalation by compressing the abdominal cavity, which increases intra-abdominal pressure and helps elevate the diaphragm.
A fundamental principle in gas physics that describes the inverse relationship between the pressure and volume of a gas at constant temperature.
The difference in pressure between two regions, which drives the movement of gases or fluids from areas of higher pressure to areas of lower pressure.
Refers to the pressure exerted by a single gas in a mixture of gases. It is a measure of how much of that gas is present and contributes to the total pressure of the mixture.
Also known as the Law of Partial Pressures, this states that the total pressure exerted by a mixture of gases is equal to the sum of the partial pressures of each individual gas in the mixture.
This describes the rate at which a gas moves across a membrane. It states that the rate of gas transfer is directly proportional to the surface area of the membrane and the difference in partial pressures across it, and inversely proportional to the thickness of the membrane.
A complex iron-containing protein found in red blood cells that is essential for oxygen transport in the body
Describes how changes in blood pH and carbon dioxide concentration influence hemoglobin’s affinity for oxygen. Specifically, a decrease in pH (more acidic) or an increase in CO₂ causes hemoglobin to release oxygen more readily—a rightward shift in the oxygen-hemoglobin dissociation curve.
The phenomenon where deoxygenated hemoglobin has a greater capacity to bind carbon dioxide (CO₂) than oxygenated hemoglobin. This effect facilitates the transport of CO₂ from tissues to the lungs.
A small, oxygen-binding protein found primarily in muscle tissue. Myoglobin stores and facilitates the transport of oxygen within muscle cells, enabling sustained energy production during periods of high metabolic demand. It consists of a single polypeptide chain and a heme group that binds oxygen molecules.
A common pulmonary function test that measures the volume and speed of air a person can inhale and exhale. It is used to assess lung function and diagnose respiratory conditions such as asthma, chronic obstructive pulmonary disease (COPD), and restrictive lung disease.
The maximum volume of air the lungs can hold after a full, deep inhalation. It includes all the air in the lungs, comprising tidal volume, inspiratory reserve volume, expiratory reserve volume, and residual volume.
The amount of air inhaled or exhaled during a normal, resting breath. It represents the baseline volume of air exchanged in the lungs without conscious effort and is typically around 500 milliliters in a healthy adult.
The total volume of air moved in and out of the lungs per minute. It is calculated as the product of tidal volume (VT) and breathing frequency (f): VE = VT × f
The number of breaths taken per minute.
The maximum amount of air a person can exhale after a full, deep inhalation. It represents the total volume of air that can be voluntarily moved in and out of the lungs and includes the inspiratory reserve volume (IRV), tidal volume (TV), and expiratory reserve volume (ERV).
The amount of air remaining in the lungs after a maximal exhalation. This air cannot be voluntarily expelled and serves to keep the alveoli open, maintaining gas exchange between breaths. RV typically accounts for about 1.2 liters in a healthy adult and is a critical component of total lung capacity (TLC).
The ratio between the amount of air reaching the alveoli (ventilation) and the amount of blood reaching the alveoli via the capillaries (perfusion).
The exercise intensity at which ventilation (breathing rate and volume) increases disproportionately to oxygen uptake (VO₂). This occurs because carbon dioxide (CO₂) production rises sharply due to increased buffering of hydrogen ions (H⁺), signaling a greater reliance on anaerobic metabolism.
The gradual increase in oxygen uptake (VO₂) during prolonged, steady-state exercise at a constant submaximal workload, typically observed after 10–15 minutes of exercise, even though the external workload remains unchanged.