A complete neuron cell, including the cell body (soma), dendrites, axon, myelin sheath, nodes of Ranvier, axon terminals, and synapse involved in the transmission of electrical impulses. Dendrites receive incoming signals, which are processed in the soma and transmitted along the axon. The myelin sheath, interrupted by nodes of Ranvier, facilitates rapid signal conduction. Axon terminals connect to other neurons or target cells via synapses. This figure represents the fundamental architecture of neurons in the vertebrate nervous system, including the brain, spinal cord, and peripheral nerves.
Learning Objectives
Describe the role of the central nervous system (CNS) in initiating and coordinating voluntary movements, including the functions of the primary motor cortex, cerebellum, and basal ganglia.
Explain the structure and function of the peripheral nervous system (PNS), particularly the motor division, and its role in transmitting impulses to skeletal muscles.
Understand the concepts of resting membrane potential, graded potentials, and action potentials, including the processes of depolarization, hyperpolarization, and the refractory periods.
Identify the steps involved in the generation and propagation of an action potential, and how it leads to muscle excitation and contraction cycling.
Discuss the mechanisms of synaptic transmission, including the roles of neurotransmitters such as acetylcholine and the processes at the neuromuscular junction.
Recognize the importance of exercise in promoting brain health, and describe the benefits of physical activity on cognitive function and overall well-being.
Summarize the pathway of force production, from CNS activation to the arrival of the action potential at the neuromuscular junction, and understand its relevance to exercise physiology and fatigue.
The Nervous System: An Overview
The nervous system is a complex network that receives millions of bits of information each minute from various sensory neurons and sensory organs. It integrates these signals to determine the necessary responses of the body. Acting as a central computer, the brain processes this information and selects appropriate responses. All functions within the human body are under the control or influence of the nervous system, which facilitates communication and coordination between different tissues and the external environment.
Generally, the nervous system, along with the endocrine system, promotes homeostasis by communicating with various tissues, organs, and systems, often without our conscious awareness. It also stores information as memories, enabling learning. Besides involuntary control, the nervous system allows volitional control of skeletal muscle locomotion. Reflexes, which can be programmed in the spinal cord, bypass higher brain centers to permit quick responses to stimuli. The nervous system transmits signals throughout the body by converting stimuli into electrical signals, or nerve impulses.
The Neuromuscular System
A single voluntary muscle contraction involves a complex pathway of events starting in the motor cortex of the brain and ending with muscle contraction cycling within a muscle fiber. This intricate process is why the nervous system and the muscular system are often grouped together and referred to as the neuromuscular system. This chapter focuses on the neural control of muscle contraction, providing an overview of the nervous system’s functions and presenting the first three steps of the pathway of force production. This pathway, designed for undergraduate students, summarizes the complex systems involved in voluntary muscle contractions. The pathway of force production includes:
Central nervous system activation of the primary motor cortex and the spinal cord
The action potential
Arrival of the action potential at the neuromuscular junction
Muscle contraction cycling
The details of processes 1-3 will be discussed in this chapter, with muscle contraction cycling and force production covered in chapter 8.
Organization of the Nervous System
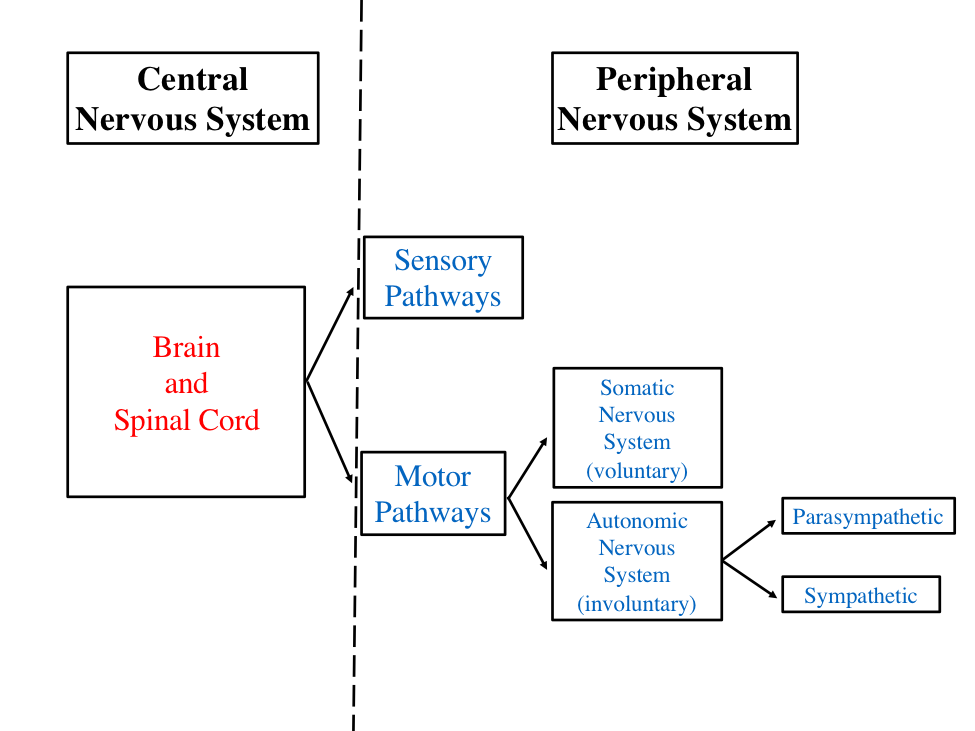
Before delving into the pathway of force production, it is essential to understand the organization of the nervous system and how it integrates and controls movement. The nervous system is divided into two anatomical systems: the central nervous system (CNS) and the peripheral nervous system (PNS). The CNS, comprising the brain and spinal cord, contains over 100 billion neurons. The PNS includes all neurons outside the CNS and is divided into sensory and motor divisions.
The sensory division of the peripheral nervous system relays impulses from receptors in the body to the CNS. Neurons that transmit messages from the body to the CNS are called afferent fibers (sensory fibers). The motor division‘s nerve cells, termed efferent fibers, carry impulses from the CNS to effector organs. An example of an efferent fiber is a motor neuron that stimulates muscle contraction in the connected muscle fiber.
Figure 7.1 Organization of the nervous system including the divisions of the central and peripheral nervous systems.
The motor division of the peripheral nervous system (PNS) is crucial in exercise physiology as it relays messages to skeletal muscles for locomotion. This division is subdivided into systems that control voluntary and involuntary actions of tissues and organs. These systems are the somatic nervous system, which pertains to the outer regions of the body, and the autonomic nervous system, also known as the visceral nervous system. The somatic nervous system is responsible for voluntary movements, whereas the autonomic nervous system governs involuntary functions. The autonomic nervous system is further divided into the sympathetic nervous system, often referred to as the “fight or flight” system, and the parasympathetic nervous system, known for “resting and digesting.” Figure 7.1 illustrates the organization of the nervous system.
Anatomy of a Neuron
The fundamental building block of the nervous system is the neuron, or nerve cell, which rapidly conveys signals over long distances to communicate and process information[1]. Neurons are specialized excitable tissues capable of transmitting electrical impulses to communicate with other neurons or tissues in the body. Anatomically, neurons share many components with typical cells, including a cell membrane, nucleus, and mitochondria. Each neuron has an enlarged cell body, known as the soma, which houses the nucleus, endoplasmic reticulum, ribosomes, and other organelles within the cytoplasm. Extending from the soma are several finger-like projections called dendrites, which make contact with neighboring neurons. At the base of the soma is the axon hillock, which integrates impulses received from other cells to determine if an action potential, or nerve transmission, will occur.
Neurons also possess a long, thin extension called the axon, which conducts impulses away from the cell body. The axon is insulated by a discontinuous myelin sheath, which facilitates rapid transmission of nerve impulses. The segments of the myelin sheath are known as Schwann cells. Near its end, the axon branches into numerous end branches, with the tips of these branches forming tiny bulbs called axon terminals. These terminals contain synaptic vesicles filled with acetylcholine, a chemical neurotransmitter. When a signal travels down the axon, acetylcholine is released into the synapse, a 20-30 nanometer gap between cells. Communication between nerve cells occurs at synapses. Synaptic clefts are too small to be observed with a light microscope, and it wasn’t until the 1950s that it was demonstrated that neurons are anatomically separate entities[2]. Figure 7.2 provides an illustration of presynaptic and postsynaptic neurons separated by a synapse, highlighting their key anatomical structures.
Figure 7.2 An illustration of a neuron and important structures. This also depicts a presynaptic cell and postsynaptic cell which are separated by a synapse.
Specialized Sensory Neurons
The central nervous system (CNS) receives constant feedback from receptors throughout the body about changes in both the internal and external environments. These receptors are sensory neurons that monitor everything from environmental sensations to the physiological status of nutrient availability. Sensory neurons include chemoreceptors, which sense changes in the internal chemical environment (e.g., H+, K+, CO2 concentrations), and baroreceptors, which sense changes in blood pressure. Given the vast number of sensory receptor variations, this discussion will focus on sensory organs relevant to exercise physiology and those responsible for body position sense. Sensory stimulation from these receptors is transmitted via sensory nerves to the spinal cord, where it can either trigger a reflex at that level or be transferred to upper regions of the spinal cord or brain.
Skeletal muscle contains several types of sensory receptors. Receptors that provide the CNS with information about the position of body parts with respect to gravity are called proprioceptors. Also known as kinesthetic receptors, proprioceptors include muscle spindles, Golgi tendon organs, and joint receptors. For the nervous system to properly control skeletal muscle movements, it must receive continuous sensory feedback about the tension building in muscles and the amount of muscle length.
The muscle spindle is the muscle’s measuring instrument for static muscle length and dynamic length changes[3]. Muscle spindles run parallel with muscle fibers and, when activated, increase the force produced by the muscle. They are composed of several thin muscle cells called intrafusal fibers, which are surrounded by connective tissue sheaths and insert into the connective tissue within the muscle fibers (extrafusal fibers). Muscle spindles have two types of sensory nerve endings: primary endings, which respond to dynamic changes in muscle length, and secondary endings, which continuously provide the CNS with information about static muscle length[4].
Figure 7.3 The structure of muscle spindles and their location in skeletal muscle.
Figure 7.4 The Golgi tendon organ. The Golgi tendon organ is located in series with the muscle and measures tension that acts as a protective mechanism for muscle.
Muscle spindles are innervated by gamma motor neurons (Figure 7.3), which stimulate the intrafusal fibers to contract simultaneously with the extrafusal fibers. When a muscle is stretched, muscle spindles activate the agonist muscle motor unit to shorten the muscle. Rapid stretching of skeletal muscles results in a reflex contraction, known as the myotatic reflex, which is monitored by the muscle spindles. The knee-jerk reflex is a classic example of the myotatic reflex, often evaluated in a physician’s office by tapping the patellar tendon with a rubber mallet. The mallet’s blow stretches the entire muscle, exciting the primary nerve endings in the muscle spindles. This stimulation activates the extrafusal fibers of the extensor muscle, the rectus femoris, resulting in the knee jerk action. Muscle spindles also help prevent collapsing or falling when muscle stretch is sensed. They detect both the stretch and the speed of the stretch.
The Golgi tendon organ (GTO) is another specialized sensory neuron that monitors tendon tension. GTOs are measuring devices for changes in muscular tension or force. Located within the tendon and in series with the extrafusal fibers (Figure 7.4), GTOs are controlled by spinal interneurons, which are subject to inhibitory influences [3]. GTOs respond more effectively to active contractile force than to passive tension produced by external pull.
The Golgi tendon organ (GTO) prevents excess force production by the muscle and also prevents muscle tearing. When activated, the GTO sends information to the spinal cord via sensory neurons. These sensory neurons, in turn, excite inhibitory neurons (IPSPs) that prevent the motor neurons from firing. This reduces muscle force production and protects the muscle against contraction-induced injury. In sports, GTOs may play an important role in performance and strength activities. It is thought that GTO influence can be gradually reduced in response to strength training, allowing an individual to produce greater muscle force by voluntarily opposing the inhibition of the GTO.
Motor Reflexes
The level of nervous system involvement in sensory input varies according to the complexity of the movement necessary to respond. Imagine a scenario where an unsuspecting person touches a hot stovetop. Within a second, the person can remove their hand from the danger without consciously thinking about the action. In the case of a reflex, the stimuli of heat and pain are received by the thermoreceptors and nociceptors in the hand, respectively, and the sensory action potentials travel to the spinal cord. Once in the spinal cord, these action potentials are integrated, and interneurons that connect the sensory and motor neurons are activated in what is referred to as a reflex arc. Once activated, action potentials in the motor neurons are sent to effectors that control the withdrawal of the hand. By the time the person is conscious of the hot surface, the reflex activity is well underway, if not completed.
A motor reflexis a rapid, unconscious means of reacting to stimuli and does not depend on higher brain centers for muscle activation. It can be thought of as a preprogrammed response, and any time the sensory nerve transmits a certain impulse, the body’s response will be instant and identical to the last. This consistency is advantageous as it eliminates the need for higher brain centers to consider options. In summary, reflexes occur when a sensory nerve sends an impulse to the spinal column. An interneuron in the spinal cord then activates motor neuron depolarization. The motor neuron immediately relays the impulse to the skeletal muscle to withdraw, bypassing higher brain centers, which would delay the reaction.
With the motor reflex activation of the agonist muscle (by EPSPs, explained later in the graded potentials section), it is also important that a simultaneous IPSP (see graded potentials) be sent to the antagonist muscle. This deactivates the antagonist muscle to prevent interference or opposition to the agonist muscle’s action. The simultaneous excitatory and inhibitory activity by the spinal cord is known as reciprocal inhibition. Along with the motor reflex, reciprocal inhibition demonstrates the spinal cord’s contribution to rapid movements. Additionally, emerging evidence suggests that reflexes play a major role in the control of voluntary movement[5][6]. Some researchers believe that additional refinement of movement occurs at the spinal cord, playing a larger role in volitional movement than initially proposed.
Somatic Alpha Motor Neurons
The somatic motor portion of the PNS is responsible for carrying neural messages from the spinal cord to the effector organs, such as skeletal muscles. The effectors of all motor actions are the skeletal muscles, whose basic properties are viscoelasticity and contractility[7]. Neural messages signal muscle contraction. The somatic neuron that innervates skeletal muscle fibers is called a motor neuron, or specifically, an alpha motor neuron. The cell body of a motor neuron is located in the spinal cord, and its axon extends to the muscle it innervates. At the muscle fiber, the axon splits into collateral branches, each innervating a single muscle fiber. Thus, one motor neuron can innervate many muscle fibers (cells). One motor neuron and all the muscle fibers it innervates are collectively known as a motor unit.
Motor units are the smallest functional entities of normal motor activity. They contain fewer than ten muscle fibers per motor neuron in the most finely adjustable muscles (e.g., extraocular muscles of the eyes), some hundreds in extremity muscles, and some thousands in very crude muscles (e.g., gluteus maximus)[8]. Mathematically, the innervation ratio can be expressed as the number of muscle fibers innervated per motor neuron (i.e., number of muscle fibers/motor neuron). When the motor neuron is activated, all the muscle fibers it innervates will simultaneously contract. Conversely, if the motor neuron ceases to contract, all the fibers it innervates will cease to contract.
Motor Unit Recruitment
The CNS can enhance muscle force production in two ways: 1) by increasing the number of activated motor units, called motor unit recruitment, and 2) by heightening the excitation frequency of individual motor units, leading to growing forces of overlapping and fusing contractions. Recruitment of additional motor units activates more muscle fibers, increasing the strength of a voluntary muscle contraction. Generally, motor units are recruited in an orderly and sequential fashion based on their size. This is called the size principle (Henneman size principle) of motor units, where recruitment begins with the smallest motor neurons and progresses to larger and larger motor neurons. For example, when a muscle is initially activated to lift a light weight, the first motor units to fire are smaller in size, as the force generation required by the muscle is low. However, if the weight is increased (heavy lifting) and the force required by the muscle increases, there will be a progressive increase in the recruitment of more and larger motor neurons in addition to the smaller motor neurons. It is also important to note that even in the strongest in vivo contractions, not all motor units of a muscle are simultaneously active[9].
The American scientist Elwood Henneman (1915-1995) discovered that the smallest motor neurons were the most easily excited, whereas the large motor neurons were the least susceptible to excitation. As the discovery of the size principle improved our knowledge of how the nervous system increases force production within the muscular system, motor units were also categorized into different types. Motor units can be divided into three classes based on their metabolic and size properties. Type S (slow) motor units are the smallest motor units and are resistant to fatigue. Type S motor units are recruited first. Type FR (fast-fatigue resistant) motor units are considered large, have a faster transmission speed of impulse, and innervate type IIa muscle fibers. Type FF (fast-fatigable) motor units have the largest motor neurons, are the last to be recruited, and innervate the largest muscle fibers (i.e., type IIx). In general, the larger the motor neuron, the faster the neural transmission speed. Additionally, motor neuron size corresponds with the size of the muscle fiber. A comparison of the characteristics of motor units is shown in Table 7.1.
Table 7.1 Characteristics of human motor unit types.
Motor Unit Type
Type S
Type FR
Type FF
Size
Small
Large
Largest
Signal Transmission Speed
Slow
Fast
Fast
Muscle Fiber Innervated
Type I
Type IIa
Type IIx
The Pathway of Force Production
The conscious desire to perform physical exercise of any intensity requires coordination of several body systems, starting in the cerebral cortex of the brain and ending with muscle force production through contraction cycling. The intricate steps in between will be discussed by system as the pathway of force production in the following sections. This pathway is extremely complex, involving the coupling of the nervous and muscular (neuromuscular) systems. It is also of particular interest in the study of fatigue, as several theories have been developed to explain decrements in performance related to neuromuscular physiology. One theory of fatigue suggests that alterations in neural control, or signaling, prevent muscle contraction. It is also thought that the CNS plays a role in most types of fatigue, perhaps limiting exercise performance as a protective mechanism[10]. Other theories propose that muscle fibers’ contractile mechanisms fail due to intracellular conditions such as acidosis. Nevertheless, it is important to realize that muscle contraction is extremely complex, and fatigue is most likely caused by multiple factors occurring at several sites along the pathway.
Central Nervous System Activation of the Primary Motor Cortex and the Spinal Cord
To comprehend how even the most basic muscle contractions are initiated, we will first consider the initial step in the pathway of force production: activation of the primary motor cortex and the spinal cord. First and foremost, the complex nature of the CNS must be investigated. In this section, a brief overview of components of the CNS will be introduced, as well as the areas of the brain that are of primary concern to exercise physiology.
The cerebrum is composed of the right and left cerebral hemispheres, connected by the corpus callosum. The cerebral cortex forms the outer portion of the cerebral hemispheres and is the conscious brain, allowing people to think, be aware of sensory stimuli, and voluntarily control movements.
Figure 7.3 A volleyball player serves the ball to the opposing team. The premotor cortex is the part of the brain where learned motor skills, like serving, are stored.
The desire and decision to perform a sport-specific skill or conscious movement of skeletal muscle begins at the primary motor cortex of the cerebrum. The primary motor cortex is the part of the brain where decisions are made about what movement to make. For example, in volleyball, if a player desires to serve the ball to the opposing team (as shown in Figure 7.3), the decision to toss the ball into the air and swing the arm to make contact with the ball is made in the primary motor cortex. Pyramidal cells are housed in the primary motor cortex, and their axons form the extrapyramidal tracts. These are known as the corticospinal tracts because the nerve processes extend from the cerebral cortex down to the spinal cord. These tracts provide the major voluntary control of skeletal muscles[11]. Learned motor skills of a repetitious nature, such as those learned in sports, are stored in the premotor cortex. The premotor cortex is located anterior to the precentral gyrus in the frontal lobe. This area of the brain can be thought of as the memory bank for skilled motor activities.
Another brain structure that is important for coordinating movement is the cerebellum. The cerebellum is located behind the brain stem, as shown in Figure 7.4. It is responsible for coordinating the timing of complex muscular activities and helps the rapid progression from one movement to the next. The cerebellum facilitates movement patterns by smoothing out the movement through corrective adjustments via the motor system. From the motor cortex, the decision to perform a movement is relayed to the cerebellum, where the desired movement is compared to the actual movement based on sensory feedback from the muscles and joints[12].
The basal ganglia (nuclei) is another area of primary concern to exercise physiology. The basal ganglia are not part of the cerebral cortex but are located in the cerebral white matter, deep within the cortex. It is thought that the basal ganglia are important in initiating movements of a sustained and repetitive nature, such as arm swinging during walking. The basal ganglia control movements such as walking and running and are also involved in maintaining posture and muscle tone. The brain stem, composed of the midbrain, the pons, and the medulla oblongata (see Figure 7.3), connects the brain and the spinal cord. Both sensory and motor neurons pass through the brain stem as they relay information between the brain and spinal cord. The spinal cord is continuous with the medulla oblongata and is composed of tracts of nerve fibers that allow two-way conduction of nerve impulses. Both afferent (towards the brain) and efferent (towards the body) nerve fibers transmit action potentials to end organs from the spinal cord. Sensory-motor integration is also assisted by specialized sensory organs within muscles as well as reflex pathways for quick responses.
Figure 7.4 The major regions of the brain associated with voluntary control of movement.
In summary, motor responses for complex movement patterns by the CNS originate in the primary motor cortex of the brain. The pathway then continues to the basal ganglia and the cerebellum, which coordinate repetitive movements and smooth out the desired movement patterns. Motor pathways (and sensory pathways) then proceed through the brain stem (midbrain, pons, medulla oblongata) and on to the spinal cord for transmission of the signal to the PNS.
Peripheral Nervous System Impulse Transmission
From the CNS, the pathway of force production continues through the motor, or efferent, division of the PNS. Cranial and spinal nerves directly innervate and carry nerve transmissions from the CNS to the skeletal muscles. The PNS contains 43 pairs of nerves. Of these, 12 pairs are cranial nerves that connect with the brain, and the remaining 31 pairs are spinal nerves that connect with the spinal cord[13]. From the brain and spinal cord, intricate networks of neurons extend to all parts of the body, including–and central to exercise and sport physiology–skeletal muscles. Signals are carried through this network by generating action potentials along the PNS until they arrive at the junction where the motor neuron and muscle meet. The details involved in nerve transmission, generation of an action potential, and arrival of the action potential at the neuromuscular junction will be discussed next.
Resting Membrane Potential
Electrical potentials exist across the membranes of virtually all cells in the body. These potentials are caused by an ion concentration difference on the two sides of the membrane. The membrane of nerve cells is polarized, meaning there is a difference in ionic charges across the membrane that creates an electric potential. Understanding the resting membrane potential of a neuron is important for understanding how an impulse, or action potential, is generated. The electrical potential difference is known as the resting membrane potential (RMP), and in most resting cells, it is measured at -70 millivolts (mV). This means that the potential inside the fiber is 70 millivolts more negative than the potential in the extracellular fluid outside the fiber[14].
The resting membrane potential is determined by the concentration of sodium (Na+), potassium (K+), and chloride (Cl–) inside and outside the cell. At rest, the inside of a neuron is negatively charged, with a large concentration of potassium inside the membrane along with negatively charged proteins that cannot cross the cell membrane. Additionally, a larger concentration of chloride and sodium exists outside the nerve cell in the extracellular space. The differences in the concentration of these ions create potential energy that can be used to transmit an action potential down the neuron axon.
Ions tend to move to establish equilibrium across the membrane as a general rule of diffusion. However, in the case of the resting membrane potential, it is necessary to maintain a difference in ion concentration across the cell membrane. This difference is maintained in two ways. First, the cell membrane is much more permeable to K+ than Na+. This means that K+ can move more freely, and some of the K+ will move to the area where it is less concentrated, outside the cell. On the other hand, Na+ cannot move inside as easily. Secondly, the membrane potential is maintained by the sodium-potassium pumps (Na+/K+ pump) located in the membrane that actively, requiring ATP, transport potassium ions in and sodium ions out. The Na+/K+ pump moves three Na+ out of the cell for every two K+ it brings in. The end result is a membrane with more positively charged ions outside the cell than inside, creating the potential difference across the membrane.
Graded Potentials, Depolarization, and Hyperpolarization
The nerve cell membrane also contains gated ion channels that act as doorways, allowing ions into and out of the nerve cell. These gates are usually closed; however, if the stimulation is potent enough, the gates open and allow more ions to move from the outside to the inside or vice versa. When ions change positions by crossing the membrane through these gates, the electrical potential will also change. Graded potentialsare localized changes in the membrane potential that occur from either depolarization or hyperpolarization. Depolarization reflects any time that the charge difference becomes more positive than the RMP of -70 mV by moving closer to zero[15]. The opposite can also occur. If the charge difference across the membrane increases, this is termed hyperpolarization, which moves the RMP to an even more negative value. Thus, hyperpolarization results in the membrane becoming more polarized.
Neurons receive signals from neighboring nerve cells that can negate an action potential or cause the action potential to be generated. The process of communication between neurons occurs through synaptic transmission. As mentioned previously, signals are transmitted at a junction called synapses via chemical messengers known as neurotransmitters. More than 50 neurotransmitters have been identified as potential candidates to serve as chemical messengers. Neurotransmitters that cause depolarization of membranes, such as acetylcholine and norepinephrine, are termed excitatory transmitters. These types of neurotransmitters bind to receptors on the target cell membrane, producing a graded potential in the dendrites and the cell body of the postsynaptic cell. Excitatory neurotransmitters released from the presynaptic cell cause excitatory postsynaptic potentials in the postsynaptic cell. Excitatory postsynaptic potentials (EPSP) cause a depolarization of the nerve cell membrane; however, the threshold is not always reached. If sufficient amounts of the excitatory neurotransmitter are released, the postsynaptic neuron can be depolarized to threshold, and an action potential will occur[16]. It is estimated that the addition of up to 50 EPSPs might be required to produce an action potential[17]. Note that not all neurotransmitters are excitatory; some can cause hyperpolarization of the membrane. Interestingly, norepinephrine can also be an inhibitory neurotransmitter depending on the receptor.
Inhibitory neurotransmitters cause the membrane to become more negative, resulting in an inhibitory postsynaptic potential (IPSP). IPSPs cause hyperpolarization of the nerve cell membrane, moving the membrane potential further away from zero. This hyperpolarization pushes the membrane potential even further from the threshold, thus resisting depolarization. The axon hillock is responsible for determining if an action potential will occur. Impulses from IPSPs and EPSPs are summed at the axon hillock, and often, more than one stimulus will be received. Whether a neuron reaches the threshold depends on the ratio of EPSPs to IPSPs received. For example, if the ratio of EPSPs to IPSPs is equal, the threshold to generate an action potential will not be reached. If the ratio of EPSPs is greater than IPSPs, the membrane potential will be pushed toward the threshold, and an action potential will occur.
Temporal summation occurs when several EPSPs from a single presynaptic neuron are received at the axon hillock over a short period. Spatial summation occurs wTemporal summationhen EPSPs are received from many presynaptic cells and are summed at several different presynaptic inputs.
Action Potential
Nerve and muscle cells are capable of generating electrochemical impulses at their membranes and can transmit signals along their membranes[18]. Neurons are known as excitable tissue because they are irritable and able to respond to a stimulus. Neurons can transmit an impulse along their axons; however, the stimulus must be large enough to create an impulse. The impulse conducted along the membrane is called an action potential. As mentioned, the RMP in most cells is -70 millivolts (mV). An action potential only occurs when a depolarization threshold of -55 mV (which makes the membrane potential more positive) is reached. Any time depolarization reaches or exceeds the threshold, an action potential will indeed result. This is known as the all-or-none principle.
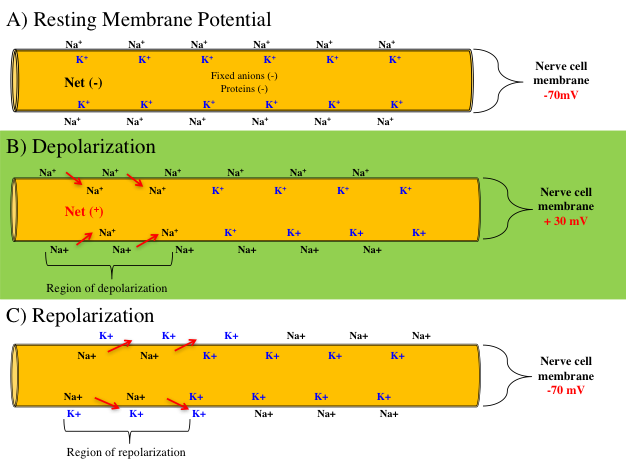
Figure 7.5 (A) The resting membrane potential is about -70 millivolts. (B) When the membrane reaches a threshold of -55 mV, sodium channels open, sodium ions rush in, and the membrane is depolarized to +30 mV. (C) When the potassium channels open, potassium diffuses outward, and the membrane is repolarized to -70 mV.
All action potentials begin as graded potentials at portions of the cell membrane[19]. When enough stimulation occurs to cause a depolarization threshold of -55 mV, an action potential will be triggered. Upon reaching the threshold, voltage-gated Na+ channels open, allowing Na+ to enter the cell rapidly. The influx of Na+ depolarizes the cell to a membrane potential of +30 mV and the action potential propagates down the axon membrane at 100 m/s (225 mph). Subsequently, the voltage-gated K+ channels open, but do so more slowly, which allows K+ to exit the cell. As K+ exits from the cell to the extracellular fluid, this causes a repolarization and returns the membrane potential to -70 mV. Also note that a slight hyperpolarization of the membrane occurs momentarily following K+ diffusion to the extracellular fluid as shown in figure 7.6. Lastly, as the membrane returns to -70 mV, the voltage-gated K+ channels close. The cell then returns Na+ and K+ back into their intracellular and extracellular resting positions via active transport of the ions by the sodium potassium (Na+K+) pump.
Figure 7.6 Voltage changes during an action potential.
An action potential occurs rapidly and substantially depolarizes the remaining portions of the membrane. Since the signal only travels away from the cell body, the depolarization will travel down the axon towards the axon terminal. When a cell is in the process of generating an action potential and a segment of an axon’s sodium gates are open, it will not be able to respond to another stimulus. This is called the absolute refractory period. When the sodium gates close and the potassium gates open (during repolarization), the segment of the axon can potentially respond to a new stimulus if the stimulus is substantially greater in magnitude. This time frame is known as the relative refractory period[20].
Arrival of the Action Potential at the Neuromuscular Junction
The nervous system’s involvement in the pathway of force production culminates when the action potential arrives at the axon terminal and interfaces with a muscle fiber. Nerve cells that extend outward from the spinal cord and transmit signals to skeletal muscles are termed alpha (α-) motor neurons. Stimulation from the nervous system via the α-motor neuron is necessary to initiate muscle excitation and contraction cycling. The axon terminal of the α-motor neuron does not physically contact the muscle fiber; instead, a small gap called the synaptic cleft separates the muscle and nerve fibers.
One α-motor neuron can innervate many muscle fibers, all of which will contract if the motor neuron depolarizes (the all-or-none principle applies here too). A single α-motor neuron and all the fibers it innervates are collectively called a motor unit. Figure 7.7 illustrates the neuromuscular junction, the site where the α-motor neuron excites the muscle fiber. The neuromuscular junction functions similarly to a synapse, except the α-motor neuron is communicating with a muscle fiber instead of a postsynaptic neuron.
Figure 7.7 Enlarged, detailed diagram of the neuromuscular junction, showing the connection between a motor neuron and a skeletal muscle fiber. The image includes the axon terminal of the motor neuron containing synaptic vesicles filled with acetylcholine, the synaptic cleft, and the motor end plate of the muscle fiber with acetylcholine receptors. The figure illustrates the process of signal transmission from the nervous system to the muscular system, highlighting neurotransmitter release, receptor binding, and initiation of muscle contraction.
When the action potential arrives at the axon terminal, a series of events occur that lead to the “coupling” of the α-motor neuron and excitation of the muscle fiber membrane. Upon arrival of the nerve impulse at the axon terminal, calcium (Ca2+) ions enter the axon terminal from the extracellular space via voltage-gated calcium channels. Calcium ions diffuse into the axon terminal and interact with synaptic vesicles filled with the neurotransmitter acetylcholine. The synaptic vesicles migrate toward the synaptic cleft and release acetylcholine into the cleft via exocytosis.
Once released, acetylcholine diffuses across the synaptic cleft and binds with acetylcholine receptors (ligand-gated cation channels) on a specialized portion of the muscle cell called the motor end plate. The motor end plate is composed of invaginated (folded to form cavities) segments of the sarcolemma. Acetylcholine receptors located on the motor end plate bind acetylcholine, causing the ligand-gated cation channels to open, allowing Na+ ions to enter the fiber and K+ ions to exit. The influx of Na+ causes the motor end plate to depolarize once the threshold has been reached. The depolarization then travels throughout the sarcolemma, down the transverse tubules, and to the sarcoplasmic reticulum of the muscle fiber. Muscle contraction cycling is then activated (see Chapter 8 for details about the transverse tubules, sarcoplasmic reticulum, and muscle contraction cycling) and is the final step in the pathway of force production.
Neurotransmission to the muscle fiber ceases when acetylcholine is removed from the synaptic cleft. This occurs when 1) acetylcholine diffuses away from the synapse, or 2) is broken down by the enzyme acetylcholine esterase into acetic acid and choline. Following acetylcholine breakdown, choline is transported back into the axon terminal for the re-synthesis of acetylcholine.
Exercise Promotes Good Brain Health
Extremely strong evidence has revealed that exercise improves brain (cognitive) function, particularly later in life. Both mental stimulation (e.g., reading) and physical exercise are interventions that can contribute to improved brain health. Numerous studies have shown that exercise targets parts of the brain involved in learning, memory, and depression, and has broad positive benefits on overall brain health. Exercise has been shown to protect against several diseases of the nervous system, such as dementia, Alzheimer’s disease, and stroke[21][22]. Regular aerobic exercise promotes brain growth factor signaling and results in:
Enhanced learning and memory
Neurogenesis, the formation of new neurons
Improved vascular function and blood flow in the brain
Attenuation of mechanisms leading to depression
Daily exercise is a simple and inexpensive way to maintain the overall function of the CNS and good brain health.
Chapter Summary
In this chapter, we explored the intricate pathway of force production, beginning with the central nervous system’s activation of the primary motor cortex and the spinal cord, and culminating in the arrival of the action potential at the neuromuscular junction. We examined the roles of various brain structures, including the cerebrum, cerebellum, basal ganglia, and brain stem, in coordinating and executing voluntary movements. The peripheral nervous system’s role in transmitting impulses to skeletal muscles was also discussed, highlighting the importance of motor neurons and the neuromuscular junction.
We investigated the mechanisms underlying resting membrane potential, graded potentials, and the generation and propagation of action potentials. The concepts of depolarization, hyperpolarization, and the refractory periods were explained to provide a comprehensive understanding of how nerve impulses are generated and transmitted. Furthermore, the critical role of neurotransmitters in synaptic transmission and the processes involved in muscle excitation and contraction cycling were discussed. The chapter also emphasized the importance of exercise in promoting good brain health, underscoring the positive effects of physical activity on cognitive function and overall well-being.
By understanding the complex interactions between the nervous and muscular systems, we gain valuable insights into the physiological processes that enable movement and the factors that can influence performance and fatigue. This foundational knowledge sets the stage for further exploration of muscle contraction cycling and force production in subsequent chapters.
Scholarly Questions
What are the two main anatomical nervous systems? How is the PNS divided?
Define the following terms: soma, dendrites, axon hillock, myelin sheath, nodes of Ranvier, and synaptic terminal.
What does depolarization mean?
What is a repolarization?
What is a graded potential?
Describe the resting membrane potential. What ions have the largest concentrations on the inside and outside of the cell?
What is the first step in the pathway of force production?
What part of the brain stores learned motor skills?
What part of the brain does movement originate?
Discuss the steps and processes of an action potential. What does it mean for a nerve cell to reach threshold?
Be able to discuss the steps and process of arrival of the action potential.
What is a motor neuron? Motor unit?
What is the principal that describes how motor units are recruited? Which motor units are recruited first?
How do IPSP’s work? EPSPs?
What is the motor end plate and where is it located?
Discuss the following statement: “Calcium has a double role in muscle contraction.”
What do GTO’s monitor? Where are they located?
What is the muscle spindle responsible for? How does it function?
Brodal P, The Central Nervous System. 2010, New York, NY: Oxford University Press. ↵
Brodal P, The Central Nervous System. 2010, New York, NY: Oxford University Press. ↵
Henatsch HD, and Langer HH, Basic Neurophysiology of Motor Skills in Sport. Int J Sports Med, 1985. 06(1): p. 2-14. ↵
Powers SK, and Howley ET, Exercise Physiology (Theory and Application to Fitness and Performance). 9th Edition ed. 2015, New York, NY: McGraw-Hill. ↵
Brodal P, The Central Nervous System. 2010, New York, NY: Oxford University Press. ↵
Henatsch HD, and Langer HH, Basic Neurophysiology of Motor Skills in Sport. Int J Sports Med, 1985. 06(1): p. 2-14. ↵
Henatsch HD, and Langer HH, Basic Neurophysiology of Motor Skills in Sport. Int J Sports Med, 1985. 06(1): p. 2-14. ↵
Henatsch HD, and Langer HH, Basic Neurophysiology of Motor Skills in Sport. Int J Sports Med, 1985. 06(1): p. 2-14. ↵
Henatsch HD, and Langer HH, Basic Neurophysiology of Motor Skills in Sport. Int J Sports Med, 1985. 06(1): p. 2-14. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Powers SK, and Howley ET, Exercise Physiology (Theory and Application to Fitness and Performance). 9th Edition ed. 2015, New York, NY: McGraw-Hill. ↵
Powers SK, and Howley ET, Exercise Physiology (Theory and Application to Fitness and Performance). 9th Edition ed. 2015, New York, NY: McGraw-Hill. ↵
Guyton AC, and Hall JE, Textbook of Medical Physiology. 11th ed. 2006, Philadelphia, PA: Elsevier Saunders. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Kenney LK, Wilmore JH, Costil DL, ed., Physiology of Sport and Exercise. 5th ed. 2012, Human Kinetics: Champaign, IL. ↵
Cotman CW, Berchtold N, Christie LA, Exercise builds brain health: key roles of growth factor cascades and inflammation. Trends Neurosci, 2007. 30(464-472). ↵
Marks BL, Katz LM, Smith JK, Exercise and the aging mind: buffing the baby boomer's body and brain. The Physician and Sportsmedicine, 2009. 37: p. 119-125. ↵
definition
The part of the nervous system consisting of the brain and spinal cord. The CNS integrates sensory information, processes signals, and coordinates responses by controlling voluntary and involuntary activities throughout the body.
The part of the nervous system located outside the brain and spinal cord, consisting of cranial nerves, spinal nerves, and associated ganglia. The PNS connects the central nervous system (CNS) to the rest of the body and is divided into the somatic nervous system (controls voluntary movements) and the autonomic nervous system (regulates involuntary functions such as heart rate and digestion).
The part of the peripheral nervous system responsible for transmitting sensory information from receptors in the body (e.g., skin, muscles, joints, and organs) to the central nervous system (CNS). It enables the perception of stimuli such as touch, temperature, pain, and body position (proprioception).
The component of the peripheral nervous system responsible for transmitting signals from the central nervous system (CNS) to muscles and glands, initiating movement and regulating bodily functions. It is divided into the somatic nervous system (controls voluntary skeletal muscle movements) and the autonomic nervous system (controls involuntary functions such as heart rate, digestion, and glandular activity).
The division of the peripheral nervous system responsible for voluntary control of skeletal muscles and the transmission of sensory information to the central nervous system (CNS). It includes motor neurons that activate muscles and sensory neurons that convey information such as touch, pain, and proprioception.
The division of the peripheral nervous system that regulates involuntary physiological functions, including heart rate, blood pressure, digestion, and respiratory rate. The ANS operates largely without conscious control and is divided into three branches: the sympathetic nervous system (prepares the body for “fight or flight”), the parasympathetic nervous system (promotes “rest and digest” functions), and the enteric nervous system (controls gastrointestinal activity).
A branch of the autonomic nervous system that prepares the body for “fight or flight” responses during stress or emergency situations. It increases heart rate, dilates airways, mobilizes energy stores, and redirects blood flow to skeletal muscles, enhancing the body’s ability to respond to perceived threats.
A branch of the autonomic nervous system that promotes “rest and digest” functions, conserving energy and maintaining homeostasis during restful states. It decreases heart rate, stimulates digestion, and supports processes such as glandular secretion and nutrient absorption.
A rapid, temporary change in the electrical membrane potential of a neuron or muscle cell that allows the transmission of signals along the cell membrane. It occurs when a stimulus causes the membrane potential to reach a threshold, triggering the opening of voltage-gated ion channels. This results in a sequence of depolarization (influx of sodium ions) and repolarization (efflux of potassium ions), followed by a return to the resting potential.
Specialized sensory receptors located in muscles, tendons, and joints that detect changes in body position, movement, and muscle tension. They provide the central nervous system with information about limb position and movement (proprioception), enabling coordination, balance, and posture control.
Specialized proprioceptive sensory receptors located within skeletal muscles that detect changes in muscle length and the rate of length change. They consist of intrafusal fibers wrapped by sensory nerve endings. When a muscle is stretched, muscle spindles send signals to the central nervous system to trigger reflexive muscle contraction (stretch reflex), helping maintain muscle tone and posture.
Specialized proprioceptive sensory receptors located at the junction between muscle fibers and tendons. They detect changes in muscle tension and force rather than length. When tension becomes too high, GTOs send inhibitory signals to the spinal cord to reduce muscle contraction, protecting muscles and tendons from potential damage. They play a key role in reflexes that regulate muscle force and maintain posture.
An automatic, involuntary response of a muscle or group of muscles to a specific stimulus, mediated by the nervous system without conscious control. Motor reflexes are typically organized through reflex arcs, which involve sensory receptors, afferent neurons, interneurons (in some cases), efferent neurons, and effectors. Examples include the stretch reflex (knee-jerk) and withdrawal reflex.
Small, temporary depolarizations of the postsynaptic membrane that occur when excitatory neurotransmitters bind to receptors, causing positively charged ions (such as Na⁺) to enter the cell. EPSPs increase the likelihood that the postsynaptic neuron will reach threshold and generate an action potential.
A temporary hyperpolarization of the postsynaptic membrane that occurs when inhibitory neurotransmitters bind to receptors, causing negatively charged ions (such as Cl⁻) to enter the cell or positively charged ions (such as K⁺) to leave. IPSPs decrease the likelihood that the postsynaptic neuron will reach threshold and generate an action potential.
Localized changes in the membrane potential of a neuron that vary in size (amplitude) depending on the strength of the stimulus. Unlike action potentials, graded potentials are not all-or-none; they can be depolarizing (excitatory) or hyperpolarizing (inhibitory) and diminish in strength as they spread from the point of origin. They occur mainly in the dendrites and cell body and play a key role in initiating action potentials when summed at the axon hillock.
A neural mechanism in which the activation of a muscle (agonist) is accompanied by the simultaneous inhibition of its opposing muscle (antagonist) to allow smooth and coordinated movement. This process is mediated by spinal interneurons within reflex arcs, such as during the stretch reflex, where contraction of the quadriceps is paired with relaxation of the hamstrings.
A functional unit of the neuromuscular system consisting of a single motor neuron and all the skeletal muscle fibers it innervates.
The number of muscle fibers controlled by a single motor neuron within a motor unit. It reflects the degree of fine motor control: muscles requiring precise movements (e.g., eye muscles) have a low innervation ratio (few fibers per neuron), while muscles generating large, powerful movements (e.g., quadriceps) have a high innervation ratio (many fibers per neuron).
A fundamental rule of motor unit recruitment stating that motor units are activated in order of increasing size of their motor neurons. Smaller motor units, which typically contain slow-twitch (fatigue-resistant) fibers, are recruited first for low-force activities. As force demands increase, progressively larger motor units with fast-twitch fibers are recruited. This ensures efficient, smooth, and energy-conserving muscle activation.
Motor units composed of a small motor neuron and slow-twitch (Type I) muscle fibers. These fibers contract slowly, generate low force, and are highly resistant to fatigue due to their rich supply of mitochondria, capillaries, and oxidative enzymes. Type S motor units are primarily recruited for sustained, low-intensity activities such as posture maintenance and endurance exercise, following the size principle.
Motor units composed of medium-sized motor neurons and fast-twitch oxidative-glycolytic (Type IIa) muscle fibers. These fibers contract quickly and produce moderate force, with greater fatigue resistance than fast fatigable units due to their mixed oxidative and glycolytic metabolism. Type FR units are recruited for activities requiring both speed and endurance, such as running or cycling at moderate intensity.
Motor units composed of large motor neurons and fast-twitch glycolytic (Type IIx or IIb) muscle fibers. These fibers contract very rapidly and generate high force but fatigue quickly because they rely primarily on anaerobic metabolism. Type FF units are recruited last, during short-duration, high-intensity activities like sprinting or heavy lifting.
Learned abilities that enable the execution of coordinated movements involving muscles and the nervous system.
Differences in electric charge across a membrane or between two points, creating a voltage that can drive the movement of ions.
The electrical potential difference across the plasma membrane of a cell when it is not actively sending signals, typically around –70 mV in neurons.
A phase in which the membrane potential of a cell becomes less negative (moves toward zero) compared to the resting membrane potential.
A change in membrane potential that makes the inside of the cell more negative than the resting membrane potential.
Temporary depolarizations of the postsynaptic membrane caused by the flow of positively charged ions (typically sodium, Na⁺) into the neuron following the activation of excitatory neurotransmitter receptors. EPSPs increase the likelihood that the postsynaptic neuron will reach the threshold to fire an action potential. They are graded potentials, meaning their strength depends on the amount of neurotransmitter released and the number of receptors activated.
The process by which multiple postsynaptic potentials (EPSPs or IPSPs) from different presynaptic neurons combine at the same time on a postsynaptic neuron. When these simultaneous inputs occur at different synapses on the dendrites or cell body, their combined effect can bring the membrane potential to threshold, triggering an action potential.
The process by which multiple postsynaptic potentials (EPSPs or IPSPs) from a single presynaptic neuron accumulate over time at the same synapse. If these signals occur in rapid succession, their combined effect can bring the postsynaptic membrane to threshold, triggering an action potential.
A fundamental property of excitable cells stating that once the threshold stimulus is reached, an action potential is generated and propagated along the membrane at full amplitude without decreasing in size. If the stimulus does not reach threshold, no action potential occurs.
The process by which the membrane potential of a cell returns to its resting negative value after depolarization. In neurons and muscle cells, repolarization occurs primarily when voltage-gated potassium (K⁺) channels open, allowing K⁺ ions to exit the cell, restoring the negative internal environment.
A specialized region of the muscle fiber’s sarcolemma (cell membrane) at the neuromuscular junction where the motor neuron communicates with the muscle.
Also known as the cross-bridge cycle, it is the repeating sequence of events that occurs during muscle contraction at the molecular level.